|
|
| |
NEUROANATOMIA
 Per quanto riguarda il midollo
spinale troviamo una porzione centrale di sostanza grigia e quando parliamo di
sostanza grigia parliamo delle regioni in cui sono particolarmente concentrati
i corpi cellulari,avvolta da un manto di sostanza bianca e quando parliamo di
sostanza bianca parliamo delle regioni dove sono particolarmente abbondanti le
fibre neuronali, che nel loro aspetto assumono anche macroscopicamente questo
colorito bianchiccio per via del fatto che la gran parte di queste è mielinizzata.Abbiamo
anche visto che la presenza dei diversi solchi che abbiamo esaminato sulla
superficie esterna del midollo spinale consente di identificare diverse regioni
nelle porzioni di sostanza bianca,dei cordoni o funicoli e anteriormente
avremo, tra la fessura mediana anteriore e la fessura laterale anteriore, che
qui non è molto evidente[1],
troviamo i cordoni anteriori; tra la fessura laterale anteriore,cioè quella da
cui esce la radice anteriore,motoria del nervo spinale e la fessura laterale
posteriore, quella in cui entra la radice posteriore sensitiva, troviamo i
cordoni laterali ma vi ricordo, che in relazione al fatto che le corna o meglio,
colonne anteriori dell'H grigia non arrivano a toccare la superficie del
midollo spinale, in realtà la separazione tra il cordone anteriore e quello
laterale è soltanto virtuale, tanto che un'importante via sensitiva che è il sistema
antero-laterale decorre in questa regione,un po' nel cordone anteriore e un
po' nel cordone laterale. Più nettamente distinti sono i cordoni posteriori i
quali decorrono tra il solco laterale posteriore e il solco mediano posteriore
e all'interno dei cordoni posteriori,in questa regione che è compresa tra i due
solchi laterali posteriori, avremo anche, soltanto per la porzione superiore
del midollo spinale,cioè dei livelli più craniali fino, di solito, a T5 come è
indicato nei testi, un cordone intermedio laterale. Questa è la topografia
della porzione di sostanza bianca del midollo spinale.
Per quanto riguarda il midollo
spinale troviamo una porzione centrale di sostanza grigia e quando parliamo di
sostanza grigia parliamo delle regioni in cui sono particolarmente concentrati
i corpi cellulari,avvolta da un manto di sostanza bianca e quando parliamo di
sostanza bianca parliamo delle regioni dove sono particolarmente abbondanti le
fibre neuronali, che nel loro aspetto assumono anche macroscopicamente questo
colorito bianchiccio per via del fatto che la gran parte di queste è mielinizzata.Abbiamo
anche visto che la presenza dei diversi solchi che abbiamo esaminato sulla
superficie esterna del midollo spinale consente di identificare diverse regioni
nelle porzioni di sostanza bianca,dei cordoni o funicoli e anteriormente
avremo, tra la fessura mediana anteriore e la fessura laterale anteriore, che
qui non è molto evidente[1],
troviamo i cordoni anteriori; tra la fessura laterale anteriore,cioè quella da
cui esce la radice anteriore,motoria del nervo spinale e la fessura laterale
posteriore, quella in cui entra la radice posteriore sensitiva, troviamo i
cordoni laterali ma vi ricordo, che in relazione al fatto che le corna o meglio,
colonne anteriori dell'H grigia non arrivano a toccare la superficie del
midollo spinale, in realtà la separazione tra il cordone anteriore e quello
laterale è soltanto virtuale, tanto che un'importante via sensitiva che è il sistema
antero-laterale decorre in questa regione,un po' nel cordone anteriore e un
po' nel cordone laterale. Più nettamente distinti sono i cordoni posteriori i
quali decorrono tra il solco laterale posteriore e il solco mediano posteriore
e all'interno dei cordoni posteriori,in questa regione che è compresa tra i due
solchi laterali posteriori, avremo anche, soltanto per la porzione superiore
del midollo spinale,cioè dei livelli più craniali fino, di solito, a T5 come è
indicato nei testi, un cordone intermedio laterale. Questa è la topografia
della porzione di sostanza bianca del midollo spinale.
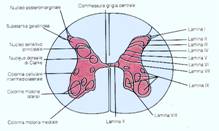
L'ultima volta che ci siamo incontrati, stavamo prendendo in esame l'organizzazione strutturale dell'H grigia, cioè delle regioni dove sono abbondanti i corpi cellulari. E qui possiamo identificare dei nuclei neuronali, intendo per tali dei raggruppamenti di neuroni i quali hanno approssimativamente la stessa morfologia e svolgono approssimativamente la stessa funzione. Quindi la prima grande classificazione funzionale dell'H grigia,prevede che le corna posteriori svolgono essenzialmente una funzione sensitiva e infatti qui arrivano le fibre che provengono dalle radici posteriori, che sono sensitive. Le fibre che provengono dalle radici posteriori sono assoni di cellule che non sono disposte all'interno del nevrasse, ma sono invece disposte nel ganglio sensitivo, con cellule a T pseudounipolari, poco prima che la radice motoria e quella sensitiva si uniscono, per cui parte un prolungamento centrifugo che andrà a raccogliere la sensibilità e un prolungamento centripeto che entrerà invece nel midollo spinale. Il corno posteriore nel complesso svolge una funzione sensitiva così come il corno anteriore svolge nel suo complesso una funzione motoria perché qui, nelle corna anteriori sono disposte delle cellule con funzione motoria, che chiamiamo motoneuroni,dai quali partiranno degli assoni che andranno a costituire la radice motoria. Nell' ambito di questa classificazione piuttosto grossolana, troviamo una regione intermedia e un cosiddetto grigio centrale che corrisponde alla sostanza grigia che è subito intorno al canale centrale del midollo e quindi abbiamo corna anteriori,corna posteriori, grigio intermedio,grigio centrale intorno al canale centrale del midollo che viene anche chiamato commensura grigia.
CLASSIFICAZIONE PER NUCLEI DEL MIDOLLO SPINALE
All'interno delle corna posteriori e anteriori identifichiamo dei nuclei. Posteriormente avremo un nucleo dorso o postero marginale,poi ancora una regione che prende il nome di sostanza gelatinosa o sostanza gelatinosa di Rolando, poi il nucleo sensitivo principale,detto così perché da lì partirà gran parte delle fibre che andrà a costituire alcune vie sensitive, poi andando verso la radice del corno posteriore, abbiamo il nucleo dorsale di Clark che avrà una funzione sensitiva molto importante, quella della sensibilità propriocettiva. Il nucleo dorsale di Clark non lo ritroveremo per tutta la lunghezza del midollo,ma soltanto a livello toraco-lombare, così come soltanto a livello toraco-lombare troveremo questa colonna cellulare intermedio-laterale che dà origine,dove è presente, ad un corno intermedio soltanto a livello toracico e quindi dei segmenti lombari,poi troveremo una colonna intermedio-mediale,la corrispondente a questa ma mediale a questa che sarà deputata alla sensibilità viscerale, poi anteriormente troveremo delle corna motorie.Questi sono i nuclei e questa è l'indicazione per nuclei del midollo spinale.
CLASSIFICAZIONE LAMINARE DI REXED
Però vi dicevo che successivamente si è identificato un altro modo di stabilire la topografia del midollo spinale e infatti nella parte destra dell'immagine[4] trovate un diverso modo di distinguere il midollo spinale. Questa è la cosiddetta classificazione laminare di Rexed. Rexed è l'anatomico che l'ha messa a punto nei primi anni '50 e questa classificazione parte dall'osservazione che tracciando dei piani approssimativamente colonnari, è possibile affettare l'H grigia, in maniera tale da identificare regioni che differiscono l'una dall'altra per la struttura delle loro cellule: è questa una citoarchitettonica spinale,vale a dire l'architettura cellulare in questo caso del midollo spinale, lo vedremo anche in altre regioni del nevrasse perché il sistema nervoso in questo caso coincide con il tessuto nervoso.Ricordate questa part 828e44i icolare terminologia per cui l'apparato nervoso in realtà viene chiamato sistema nervoso per cui si dice che è costituito da un unico tessuto,cioè quello nervoso. Però tutto sommato, dire che è un unico tessuto potrebbe quasi indurre in equivoco,perché noi chiamiamo sì il tessuto nervoso unico tessuto,ma esso è costituito da popolazioni cellulari che sono molto diverse tra di loro.Distinguiamo non soltanto le cellule neuronali dalle diverse popolazioni di cellule gliali,ma anche le diverse popolazioni di cellule neuronali;a differenza di quanto accade in altri tessuti,per esempio il tessuto osseo dove le cellule sono tutte uguali, svolgono tutte la stessa funzione,nel tessuto nervoso, abbiamo popolazioni cellulari che sono morfologicamente molto diverse tra di loro,svolgono funzioni molto diverse tra di loro,e che soprattutto,e questa è poi la base di tutte le altre caratteristiche,hanno una composizione chimica e un metabolismo molto diverso tra di loro. Dire neurone non significa identificare un tipo di cellula che svolge esattamente quella funzione se non quella caratteristica generale, di cui parlavo la volta scorsa,che peraltro è comune alle cellule endocrine,che sono in grado di produrre e di liberare nell'ambiente esterno molecole che avranno su un'altra cellula un effetto di regolazione; ma nell'ambito di questa definizione molto generale, troviamo popolazioni molto eterogenee all'interno delle diverse sottopopolazioni neuronali,gruppi di cellule che sono morfologicamente diverse tra di loro,che hanno un metabolismo molto distinto, che hanno dei componenti chimici all'interno molto distinti, che hanno delle connessioni ben distinte tra di loro,e che quindi come esito di tutto questo, svolgono delle funzioni ben diverse tra di loro. Questo è un concetto, se si vuole ovvio,ma che dobbiamo tenere sempre ben presente,perché quando parliamo di neuroni utilizziamo una terminologia molto generale,mentre poi dovremo andare ad identificare le individuali popolazioni neuronali. Sulla base di questo assunto che non è dogmatico, né un dato verificato sperimentalmente,è stata impostata quest'altra classificazione del midollo spinale che nasce proprio dal fatto che se uno affetta più o meno il midollo spinale secondo piani coronali,si imbatte in gruppi di neuroni i quali sono strutturalmente tutti uguali o molto simili tra di loro,nell'ambito di ognuna di queste fette,di queste lamine,per usare il termine più correttamente impiegato e che però sono o molto o poco diverse dalle cellule delle lamine vicine o di altre lamine lontane; cioè se uno esamina al microscopio con molta attenzione vedrà che ci sono delle lamine in cui le cellule sono più o meno tutte uguali, poi ad un certo punto si passa ad un'altra lamina in cui le cellule sono diverse da quelle della lamina affianco,ma sono tra di loro ancora più o meno tutte uguali. Ciò ha dato origine alla classificazione laminare di Rexed, per cui le corna anteriori, posteriori e regione centrale del midollo spinale sono state suddivise in dieci lamine che hanno una numerazione crescente andando da dorsale verso ventrale,da uno a nove e la decima è quella che troviamo tutt'intorno al canale centrale del midollo,quindi dove c'è la commensura grigia,cioè la regione che mette in comunicazione la porzione destra con la porzione sinistra dell'H grigia, questa (la commessura grigia) corrisponde alla lamina decima e approssimativamente possiamo trovare una certa corrispondenza tra la organizzazione in nuclei, che storicamente precede questa di Rexed,ma che tutt'ora troverete di frequente sui testi di neuroanatomia,e la classificazione laminare di Rexed.
Per cui quello che abbiamo definito nucleo postero marginale o dorso marginale,una specie di cappuccio estremo che si trova all'estremità posteriore dell'H grigia corrisponde alla lamina numero uno.
La sostanza gelatinosa o sostanza gelatinosa di Rolando che ha questo nome per l'aspetto gelatinoso che macroscopicamente essa assume, corrisponde alla lamina seconda. Il nucleo proprio sensitivo prende invece la lamina terza, quarta e quinta e anche qui troverete delle leggere differenze nei testi, ma nella maggior parte di essi corrisponderà alla lamina terza, quarta,quinta, cioè il nucleo sensitivo proprio in realtà corrisponde più o meno all'insieme di tre lamine,quindi è un nucleo molto importante. Invece nella lamina settima, troveremo sia il nucleo dorsale di Clark sia il nucleo intermedio mediale che il nucleo intermedio laterale,questi saranno tutti compresi in questa regione piuttosto intermedia che è la lamina settima.
I motoneuroni saranno tutti presenti tra la lamina ottava e nona, abbiamo raggiunto una numerazione più alta, ci troviamo appunto nelle corna anteriori del midollo.
Quindi se vogliamo riassumere.la classificazione di Rexed è tale per cui l'H grigia viene distinta in lamine, le quali vengono connotate con una numerazione che va da uno a dieci, in maniera tale che la numerazione più bassa corrisponde alle corna posteriori, la numerazione più alta alle corna posteriori e in realtà poi la lamina decima non la troviamo in corrispondenza delle corna anteriori ma in corrispondenza della commensura grigia. C'è anche una certa corrispondenza tra divisione laminare e la suddivisione in nuclei più tradizionale. Nei diversi testi,troverete o entrambe le identificazioni o una delle due.
LE DUE SENSIBILITA' DELLE RADICI POSTERIORI DEL MIDOLLO
Adesso che abbiamo un inquadramento generale sia macroscopico che strutturale del midollo spinale,vediamo quali sono le connessioni che per ora vediamo in maniera molto grossolana.
La prima cosa, procedendo da dietro in avanti, cominciamo ad esaminare le regioni sensitive poi vedremo quelle motorie. Nelle regioni sensitive troviamo le radici posteriori. Abbiamo visto questi che sono assoni che provengono da cellule a T che si trovano nel ganglio sensitivo spinale.
Andando più nei dettagli vediamo che tipo di sensibilità viene portata nelle radici sensitive del midollo. Verranno portate principalmente sensibilità di tipo dolorifico e termico.Nella porzione laterale della radice, noi vedremo che arrivano fibre che portano la sensibilità dolorifica e termica,nella regione mediale della radice quindi quella più vicina al piano sagittale mediano, troviamo fibre che portano la sensibilità tattile e la sensibilità propriocettiva. Quest'ultima proviene dai muscoli dei tendini e in ogni momento ci informa dello stato di contrazione, della posizione di un fascio muscolare, ci dà quello che si definisce in termini più conversativi il cosiddetto senso di noi nello spazio, cioè noi in ogni momento sappiamo bene anche senza guardarlo se un nostro arto è disteso oppure è flesso perchè abbiamo informazioni che provengono dai muscoli e dai nervi.
Torniamo per il momento su
queste due forme di sensibilità che seguiranno delle vie molto diverse tra di
loro all'interno del nevrasse. La sensibilità che noi seguiremo per prima e che
ci guiderà lungo la risalita del nevrasse sarà la sensibilità tattile e
propriocettiva perché queste due forme di sensibilità andranno insieme lungo il
nevrasse e vedremo poi in che modo si comportano le fibre e le cellule che
portano la sensibilità dolorifica e termica. Vorrei però mettere in evidenza
sin da ora il modo in cui interagiscono le fibre che portano questi due diversi
tipi di sensibilità una volta che entrano nel midollo spinale. Queste fibre che
arrivano all'interno del midollo spinale, avranno dei destini diversi, nel
senso che le fibre della sensibilità dolorifica e termica andranno direttamente
a raggiungere cellule prevalentemente in quel nucleo sensitivo principale e lì
contrarranno subito sinapsi con queste cellule che poi seguiranno il decorso
che vedremo ins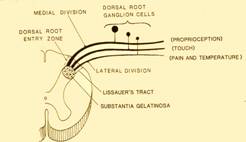 ieme. Le fibre invece della
sensibilità tattile e propriocettiva entreranno nel midollo spinale però la
fibra principale si disporrà qui[5]
all'interno del cordone posteriore del midollo per salire su e andare a
contrarre sinapsi più importante un pochino più in alto.
ieme. Le fibre invece della
sensibilità tattile e propriocettiva entreranno nel midollo spinale però la
fibra principale si disporrà qui[5]
all'interno del cordone posteriore del midollo per salire su e andare a
contrarre sinapsi più importante un pochino più in alto.
Però anche le fibre della sensibilità tattile e propriocettiva secondo una sorta di regola generale che abbiamo già visto a proposito del fascicolo proprio, mandano dei rami collaterali che entrano poi nel corno posteriore del midollo e le fibre che portano la sensibilità dolorifica e termica e i rami collaterali delle fibre che portano la sensibilità tattile interagiscono tra di loro a livello del corno posteriore del midollo in modo tale che le fibre della sensibilità tattile interferiscono con la sensibilità dolorifica. Tutto questo lo riprenderemo più in la quando sarà più chiaro per voi ma credo sia utile fin da ora anticiparvi questo concetto perché subito ci introduce al ruolo e all'importanza delle cellule del secondo tipo di Golgi, cellule piccole che hanno gli assoni che non vanno molto lontano rispetto al territorio di competenza dei dendriti, i cosiddetti interneuroni che svolgono ruoli importanti di modulazione, vi ho anticipato il concetto di generatore centrale di schema motorio,un concetto piuttosto generico per la motilità. In questo caso vediamo un altro modo che è stato studiato in maniera più dettagliata ,anche perché più semplice, e che è più utile per comprendere l'importanza di queste popolazioni cellulari.
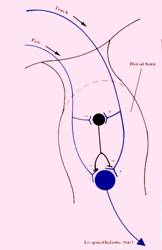 Questo sarà il corno
posteriore del midollo[6],
questa sarà una cellula disposta nel nucleo sensitivo principale,quello che
prende la lamina terza, quarta e quinta e da queste cellule partiranno delle
fibre che andranno su a portare la sensibilità dolorifica e termica. Queste
sono quindi cellule importantissime per la sensibilità dolorifica e termica e
da lì parte il passaggio successivo della sensibilità dolorifica e termica. A
queste cellule che sono nel nucleo principale arrivano le fibre dalla radice
posteriore del midollo disposte lateralmente e questa particolare disposizione
che ha un senso, per cui nella porzione laterale della radice posteriore del midollo
troviamo le fibre che portano la sensibilità dolorifica e termica mentre nella
porzione mediale della radice posteriore del nervo spinale passano le fibre che
portano la sensibilità tattile e propriocettiva, quindi a questa cellula del
nucleo sensitivo principale arrivano fibre che stimolano le cellule sensitive
principali che provengono dalla periferia per portare avanti la sensibilità
dolorifica e termica.
Questo sarà il corno
posteriore del midollo[6],
questa sarà una cellula disposta nel nucleo sensitivo principale,quello che
prende la lamina terza, quarta e quinta e da queste cellule partiranno delle
fibre che andranno su a portare la sensibilità dolorifica e termica. Queste
sono quindi cellule importantissime per la sensibilità dolorifica e termica e
da lì parte il passaggio successivo della sensibilità dolorifica e termica. A
queste cellule che sono nel nucleo principale arrivano le fibre dalla radice
posteriore del midollo disposte lateralmente e questa particolare disposizione
che ha un senso, per cui nella porzione laterale della radice posteriore del midollo
troviamo le fibre che portano la sensibilità dolorifica e termica mentre nella
porzione mediale della radice posteriore del nervo spinale passano le fibre che
portano la sensibilità tattile e propriocettiva, quindi a questa cellula del
nucleo sensitivo principale arrivano fibre che stimolano le cellule sensitive
principali che provengono dalla periferia per portare avanti la sensibilità
dolorifica e termica.
Voi vedete qui anche questa cellulina nera[7],che è un interneurone disposto prevalentemente nella sostanza gelatinosa di Rolando nella lamina seconda ed è un interneurone inibitorio. Esso manda le sue fibre alla cellula del nucleo sensitivo principale e quando questo interneurone inibitorio è attivo,inibisce la cellula del nucleo sensitivo principale e quindi la sensibilità dolorifica e termica non passa, perché la cellula principale è inibita. Dobbiamo abituarci all'algebra di più e meno per cui ci saranno cellule inibitorie che vanno a inibire cellule eccitatorie. Questo interneurone inibitorio se è attivo,inibisce la cellula del nucleo sensitivo principale. L'interneurone inibitorio che è in grado di inibire la cellula del nucleo sensitivo principale, a sua volta è raggiunto da rami collaterali delle due diverse componenti della informazione sensitiva:in primo luogo dai rami collaterali delle fibre che portano la stessa sensibilità dolorifica e termica, vale a dire: le stesse fibre che portano la sensibilità dolorifica e termica e che vanno quindi a stimolare le cellule del nucleo sensitivo principale affinchè la sensibilità dolorifica e termica proceda, mandano dei rami collaterali inibitori agli interneuroni che a loro volta sono inibitori, per bloccarli, come se volessero garantire il buon esito della propria funzione, perché loro da un lato vanno a stimolare la cellula della sensibilità dolorifica e termica, dall'altro vanno ad inibire le cellule inibitorie,cioè evitano il rischio che queste cellule inibitorie vadano ad inibire le cellule del nucleo sensitivo principale.
D'altro canto però sempre nella stessa radice sensitiva della porzione mediale del midollo spinale arrivano le fibre della sensibilità tattile e propriocettiva. Anche queste fibre mandano dei rami agli interneuroni inibitori che sono disposti nella sostanza gelatinosa di Rolando,ma questi rami che arrivano dalla sensibilità tattile e propriocettiva stimolano gli interneuroni inibitori, svolgono quindi un ruolo opposto rispetto alle fibre che portano la sensibilità dolorifica. Quindi gli interneuroni inibitori sono da un lato inibiti dalle cellule che portano la sensibilità dolorifica, e dall'altro sono stimolati dalle cellule che portano la sensibilità tattile e propriocettiva. Queste cellule (gli interneuroni)svolgono un ruolo chiave perché se queste cellule sono inibite allora la sensibilità dolorifica e termica passa,se queste cellule,che sono inibitorie, sono stimolate allora la sensibilità dolorifica e termica non passa attraverso le cellule del nucleo sensitivo principale.
Vengono chiamate cellule cancello: (sempre gli interneuroni)possono tenere il cancello aperto se queste cellule sono inibite,il cancello si chiuderà invece se queste cellule sono stimolate. Quindi in ogni momento, il passaggio o meno della sensibilità dolorifica in presenza di uno stimolo dolorifico dipenderà dalla prevalenza a livello di questi interneuroni della sostanza gelatinosa di Rolando, cioè di queste cellule cancello,dello stimolo dolorifico contro un ipotetico stimolo tattile e propriocettivo. Su questo è basata l'esperienza che ognuno di noi ha fatto tante volte nella vita,e cioè che appena abbiamo sensibilità dolorifica in qualunque parte del nostro corpo,siamo subito portati a toccare quella parte del nostro corpo perché abbiamo imparato che toccando quella parte del nostro corpo la sensibilità dolorifica si riduce perché in quel momento aumenta lo stimolo da parte delle fibre tattili che vanno ad attivare le cellule inibitorie. Su questo torneremo perché la modulazione della sensibilità dolorifica è un fenomeno più complesso di questo, però la base fondamentale della modulazione della sensibilità dolorifica consiste proprio nella presenza di queste cellule modulatorie che si trovano nella sostanza gelatinosa di Rolando le quali si comportano in maniera tale che se queste cellule inibitorie sono attive non passa la sensibilità dolorifica,il cancello è chiuso; se invece queste cellule inibitorie sono inattive,cioè se prevale lo stimolo dolorifico,le cellule inibitorie sono inattive e il cancello è aperto, quindi la sensibilità dolorifica passa.
Questo sistema è utile per comprendere il ruolo chiave svolto dagli interneuroni nell'economia generale del sistema nervoso.
Abbiamo visto le connessioni più importanti che troviamo a livello del corno posteriore. Abbiamo visto che arrivano diverse componenti di fibre con funzioni distinte disposte in regioni diverse e questo poi è in relazione al destino che avranno le diverse fibre,abbiamo visto il modo in cui interagiscono tra di loro e in pratica gli interneuroni che troviamo nel corno posteriore.
Nel corno anteriore del midollo invece troviamo dei motoneuroni, cellule che inviano fibre ai diversi fasci muscolari . Queste cellule che inviano fibre che andranno a costituire la radice anteriore del nervo spinale, saranno naturalmente,le loro fibre escono addirittura dal midollo spinale, secondo la classificazione di Golgi, cellule del primo tipo di Golgi perché inviano gli assoni molto lontano. Molto diverso dalla organizzazione strutturale degli interneuroni di cui sopra.Questi assoni usciranno dal midollo spinale e andranno a raggiungere le diverse popolazioni muscolari. Vedremo poi come sono collocate queste cellule nella gerarchia generale delle vie motorie.
Per ora esaminiamo invece la cosiddetta SOMATOTOPIA MOTORIA SPINALE. Presumo che già abbiate questo concetto di somatotopia,vale a dire la identificazione a livello delle diverse porzioni del sistema nervoso,qui siamo nel midollo spinale quindi abbiamo una somatotopia spinale, che ci informa della corrispondenza tra le diverse popolazioni neuronali, che equivale a dire diversi nuclei, e le diverse parti del nostro corpo che essi innervano. In questo caso è una somatotopia motoria,nel senso che esaminiamo ora la corrispondenza di un gruppo di cellule nervose motorie,cioè di motoneuroni e di specifici fasci muscolari.
Noi sappiamo a livello spinale precisamente dove sono i motoneuroni che vanno ad innervare dei precisi fasci muscolari. Questa è una somatotopia,cioè corrispondenza tra le diverse parti del corpo,in diversi luoghi del sistema nervoso,motoria perché stiamo parlando di una somatotopia, una corrispondenza tra motoneuroni e fasci muscolari;incontreremo anche delle somatotopie sensitive, cioè la corrispondenza tra diverse popolazioni di neuroni e di diverse parti superficiali del nostro corpo da cui queste ricevono informazioni sensitive;spinale perché siamo a livello del midollo spinale;incontreremo poi delle somatotopie o motorie o sensitive a livello delle diverse regioni del nevrasse dove risaliamo.
Nelle corna anteriori del midollo distinguiamo fondamentalmente due gruppi di nuclei neuronali: un nucleo disposto medialmente e un nucleo disposto lateralmente. Questi due nuclei hanno funzioni diverse, hanno una diversa somatotopia nel senso che i nuclei disposti medialmente innerveranno la muscolatura assile con funzione anti-gravitaria,che controlla la postura,quello stato di contrazione più o meno diffusa dei nostri diversi fasci muscolari che fanno sì che possiamo mantenere la stazione eretta contro la forza di gravità e questa condizione di contrazione molto limitata di alcuni dei nostri fasci muscolari è il punto di partenza di tutti i nostri movimenti volontari.La postura è controllata da popolazioni di motoneuroni che sono disposti medialmente nelle corna anteriori. Lateralmente invece saranno disposti gruppi di motoneuroni che andranno ad innervare fondamentalmente la muscolatura degli arti superiori e inferiori. Quindi questa colonna laterale sarà particolarmente rappresentata a livello dei rigonfiamenti cervicali e lombari. Il midollo spinale ha una conformazione esterna tale che a livello cervicale e lombare ritroviamo due rigonfiamenti perché lì sono presenti oltre alle cellule che servono per l'innervazione motoria e sensitiva di tutto il tronco,anche quelle che servono per l'innervazione motoria e sensitiva degli arti:quello cervicale per gli arti superiori,quello lombare per gli arti inferiori. Quindi questa colonna laterale del corno del midollo sono proprio le cellule responsabili dei movimenti volontari, quindi prevalentemente in relazione ai gruppi muscolari che troviamo negli arti superiori e inferiori.
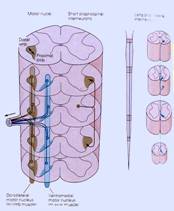
Questa somatotopia è tale appunto che mano a mano che ci allontaniamo nel corno anteriore del midollo e a mano a mano che ci allontaniamo dal piano sagittale mediano,ci allontaniamo, nei motoneuroni, sempre più anche dal tronco,in maniera tale che in posizione mediale troveremo la muscolatura che innerverà la muscolatura assile, quella del tronco e poi mano a mano, qui siamo in presenza del rigonfiamento cervicale[8],cioè dell'innervazione dell'arto superiore, troveremo dapprima i muscoli della spalla, poi del braccio,dell'avambraccio e infine della mano, cioè sempre più lateralmente sono disposti i motoneuroni che innervano popolazioni muscolari più distali, cioè più lontani dal tronco.Ricordate questo concetto di prossimale e distale riguardo alla distanza dal tronco.
Quindi la somatotopia motoria spinale è tale che i motoneuroni disposti medialmente innervano la muscolatura assile anti-gravitaria e poi a mano a mano che ci allontaniamo dal piano sagittale mediano troviamo motoneuroni che innerveranno fasci muscolari degli arti che sono sempre più lontani dal tronco, dall'attacco dell'arto.Questa suddivisione in queste due grandi colonne mediale e laterale avrà una grande importanza e la esamineremo quando troveremo le vie che controllano le vie motorie, perché troveremo delle vie di controllo motorio che specificamente saranno deputate al controllo della postura e che quindi andranno a raggiungere questi motoneuroni disposti medialmente, mentre ci sono altre vie di controllo motorio che saranno specificamente deputate alla motilità volontaria e che andranno quindi a raggiungere la colonna disposta lateralmente. Non è un aspetto semplicemente descrittivo ma ha una certa importanza e lo ritroveremo.
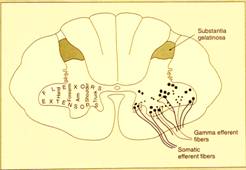
In questa immagine[9]nella parte sinistra,c'è quest'altro aspetto relativo alla somatotopia che ha grande importanza e cioè, fermo restando che i motoneuroni disposti medialmente sono quelli che innervano il tronco e che a mano a mano che ci spostiamo lateralmente troviamo motoneuroni che innervano dapprima la spalla e così via fino ad arrivare alla mano,nell'ambito di questo, i motoneuroni disposti anteriormente,anche questo è noto, innervano i muscoli estensori,mentre i motoneuroni disposti posteriormente innervano i muscoli flessori.
La parte destra dell'immagine invece ci introduce ad un altro concetto di cui parliamo molto brevemente perché
 occorre parlarne dal punto
di vista della struttura che sarà un concetto molto importante per voi in
fisiologia proprio quando studierete i meccanismi fisiologici che controllano l
occorre parlarne dal punto
di vista della struttura che sarà un concetto molto importante per voi in
fisiologia proprio quando studierete i meccanismi fisiologici che controllano l
a postura di cui molto grossolanamente vi ho dato un cenno, ma che è un capitolo molto importante nello studio della fisiologia del sistema nervoso .Questo aspetto sarà molto importante quando studierete il controllo della postura.
Sulla parte destra dell'immagine notiamo queste che sono tutte le diverse popolazioni, diversi nuclei di motoneuroni. Vedete che poi qui[10] troviamo dei puntini più densi e grossi e degli altri puntini più piccoli. Sia dagli uni che dagli altri partono delle linee.I puntini naturalmente rappresentano il corpo cellulare dei motoneuroni,le linee rappresentano il decorso di assoni che partono dai motoneuroni. Saranno tutte cellule del primo tipo di Golgi e anche cellule radicolari del primo tipo di Golgi. Ricordiamo la differenza tra cellule del primo tipo che inviano i loro assoni molto lontano dal territorio di competenza della propria arborizzazione dendritica e poi fondamentalmente distingueremo delle cellule radicolari che inviano gli assoni a costituire le radici dei nervi, le radici motorie e delle cellule funicolari che ugualmente inviano gli assoni molto lontano ma rimarranno nel nevrasse e andranno a raggiungere o altri centri in basso o in alto nel nevrasse oppure, nelle grandi vie sensitive,andranno necessariamente verso l'alto.
MOTONEURONI E
Comunque nell'ambito di questi che sono tutti motoneuroni del primo tipo di Golgi radicolari[12],vediamo qui fondamentalmente due tipi di cellule,due popolazioni: una più grande e una più piccola.
Quelle più grandi rappresentano i cosiddetti motoneuroni . Sono quelli che effettivamente vanno a raggiungere i diversi fasci muscolari e quelli il cui eccitamento determinerà la contrazione del muscolo.
Quelli più piccoli invece sono i motoneuroni cui ovviamente,ora qui dobbiamo accennare parlando noi della struttura del sistema nervoso ma che saranno molto importanti nello studio della fisiologia e di cui avete già notizia dalla istologia. Essi raggiungono in realtà una struttura sensitiva proprio per quella sensibilità propriocettiva,cioè quella sensibilità che ci dà informazioni a proposito delle condizioni di contrazione o rilasciamento dei muscoli, quella che ci dà il senso di noi nello spazio, queste fibre che sono fibre motorie più piccole, vanno a raggiungere degli organi sensitivi, i fusi neuromuscolari.
Essi sono strutture interposte nei diversi fasci muscolari che troviamo nel nostro organismo e vengono stimolati quando il muscolo si distende. Hanno al centro una regione che riceve fibre sensitive e fin qui siamo assolutamente nell'ambito della sensibilità, al centro dei quali abbiamo delle fibre sensitive. Quando il muscolo si stende,queste fibre sensitive sono eccitate e informano il nostro sistema nervoso centrale che il muscolo in quel momento è disteso.Questo è parte della sensibilità propriocettiva.
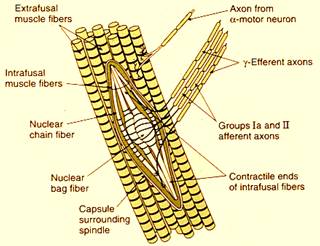 La porzione centrale del
fuso neuromuscolare è la porzione in cui arrivano queste informazioni
sensitive, queste fibre afferenti. Ad entrambe le estremità di questa porzione
centrale, questa è raggiunta da piccole fibre muscolari che vengono dette fibre
intrafusali . Queste fibre muscolari intrafusali sono raggiunte proprio
da fibre di questi motoneuroni quelli più piccoli.
La porzione centrale del
fuso neuromuscolare è la porzione in cui arrivano queste informazioni
sensitive, queste fibre afferenti. Ad entrambe le estremità di questa porzione
centrale, questa è raggiunta da piccole fibre muscolari che vengono dette fibre
intrafusali . Queste fibre muscolari intrafusali sono raggiunte proprio
da fibre di questi motoneuroni quelli più piccoli.
Ricapitolando: per ogni fascio muscolare,abbiamo i motoneuroni . che vanno a raggiungere le vere e proprie fibrocellule muscolari, quelle che determinano la contrazione del muscolo, le cosiddette fibre extrafusali, che sono le vere e proprie cellule del muscolo,le fibrocellule muscolari,che determinano con il loro eccitamento, la contrazione del muscolo. Quindi al muscolo arrivano delle fibre extrafusali,che sono innervate dai motoneuroni quelli più grandi. Contemporaneamente allo stesso muscolo arrivano delle più piccole fibre nervose dai motoneuroni ,quelli più piccolini che per lo stesso muscolo,quindi sempre nell'ambito della stessa somatotopia,questi motoneuroni non raggiungono le fibre extrafusali, le vere e proprie fibre muscolari ma raggiungono queste più piccole fibre muscolari che si trovano all'interno del fuso neuromuscolare,le cosiddette fibre intrafusali, le quali con la loro contrazione che sarà determinata dallo stimolo che proviene dalle fibre , quando il muscolo si contrae, quindi quando sono in azione le fibre , queste determineranno una contrazione del muscolo un accorciamento. In questo caso quindi si riduce la stimolazione perché il fuso viene stimolato quando viene disteso il muscolo, in questo caso quindi si riduce la stimolazione del fuso neuromuscolare. Se contemporaneamente, come accade, entrano in funzione anche le fibre , queste determineranno una contrazione delle fibre intrafusali che tireranno dalle due parti la parte sensibile del fuso neuromuscolare e in qualche modo mimeranno una distensione del muscolo anche quando questo è contratto. Tutto questo fa sì che il fuso neuromuscolare mantenga sempre una piccola condizione di eccitamento che svolge un ruolo importante nel generale controllo motorio.
Per cui dal punto di vista della struttura noi dobbiamo sapere che per ogni parte di fascio muscolare nell'ambito di ognuna di queste regioni la cui somatotopia stiamo organizzando e abbiamo visto qual è la somatotopia: medialmente la muscolatura assile e man mano si va verso la mano,verso le estremità degli arti,per ognuno di questi nuclei di motoneuroni che vanno verso i muscoli, non soltanto abbiamo i veri e propri motoneuroni che innervano il muscolo cioè i motoneuroni che vanno ad innervare le fibre muscolari che determineranno poi la contrazione del muscolo nel suo complesso, ma nella stessa regione contemporaneamente vanno anche delle fibre di altri motoneuroni più piccoli che all'interno dello stesso muscolo non raggiungeranno le grandi fibre extrafusali ma raggiungeranno invece le piccole fibre intrafusali nel fuso neuromuscolare, ne determineranno la contrazione contemporaneamente alla contrazione delle fibre extrafusali e questo farà sì che sarà sempre un po' distesa, quindi sempre un po' eccitata questa regione sensitiva centrale del fuso neuromuscolare che risponde ad una sua distensione. Quando non c'è la distensione vera e propria del muscolo, c'è però sempre una piccola distensione che proviene dalla contrazione delle fibre intrafusali che per così dire tirano da una parte e dall'altra la porzione sensibile che troviamo al centro del fuso neuromuscolare.
Questo dal punto di vista funzionale è una iper semplificazione, però dal punto di vista della struttura,che è ciò che ci interessa,teniamo conto che per ogni fascio muscolare, per ogni regione la cui somatotopia abbiamo esaminato,partono i motoneuroni che raggiungono le fibre extrafusali,ma anche con la stessa destinazione i motoneuroni che raggiungono invece le fibre intrafusali.
CELLULE DI RENSHAW
A proposito di popolazioni che troviamo nelle corna anteriori del midollo,un'altra popolazione molto importante, il cui ruolo verrà di volta in volta esaminato, è quella delle cosiddette cellule di Renshaw,che svolgono anch'esse un ruolo di modulazione molto importante e prendono il nome da chi le ha studiate. Sono cellule inibitorie, cellule a glicina.[13]
Vedete qual è l'organizzazione generale :un motoneurone che sarà un motoneurone ,invia la sua fibra a costituire la radice anteriore del midollo spinale quella motoria, però anche in questo caso come abbiamo visto per le fibre sensitive,manda un piccolo ramo collaterale subito dopo l'origine dell'assone, che va ad eccitare questa cellula schematizzata in nero che è la cellula di Renshaw che è una cellula inibitoria a glicina.
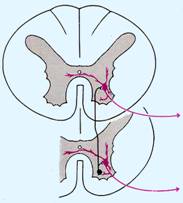 Gli assoni delle cellule di
Renshaw in primo luogo ritornano sullo stesso motoneurone da cui sono stimolati
cioè determinano quella che viene chiamata INIBIZIONE RICORRENTE cioè
controllano,limitano l'eccitamento del neurone da cui sono innervati. E' una
sorta di auto-limitazione del neurone nella sua stimolazione, nel momento in
cui parte l'eccitamento da questo neurone,esso manda un ramo collaterale
eccitatorio su una cellula inibitoria che è la cellula di Renshaw che a sua
volta manda un ramo che ritorna su quello stesso neurone, quindi limita
l'eccitamento di quel neurone,perché la cellula di Renshaw che è inibitoria
andrà ad inibire lo stesso motoneurone da cui essa stessa è stata stimolata e
oltre che lo stesso motoneurone, andrà ad inibire i motoneuroni circonvicini
dei diversi segmenti che svolgono una funzione analoga. E' una sorta di
controllo interno, di auto-limitazione dell'eccitamento neuronale. Quest'altra
popolazione neuronale che troviamo nelle corna anteriori ha un ruolo importante
nell'auto-controllo a livello spinale dell'eccitamento dei motoneuroni.
Gli assoni delle cellule di
Renshaw in primo luogo ritornano sullo stesso motoneurone da cui sono stimolati
cioè determinano quella che viene chiamata INIBIZIONE RICORRENTE cioè
controllano,limitano l'eccitamento del neurone da cui sono innervati. E' una
sorta di auto-limitazione del neurone nella sua stimolazione, nel momento in
cui parte l'eccitamento da questo neurone,esso manda un ramo collaterale
eccitatorio su una cellula inibitoria che è la cellula di Renshaw che a sua
volta manda un ramo che ritorna su quello stesso neurone, quindi limita
l'eccitamento di quel neurone,perché la cellula di Renshaw che è inibitoria
andrà ad inibire lo stesso motoneurone da cui essa stessa è stata stimolata e
oltre che lo stesso motoneurone, andrà ad inibire i motoneuroni circonvicini
dei diversi segmenti che svolgono una funzione analoga. E' una sorta di
controllo interno, di auto-limitazione dell'eccitamento neuronale. Quest'altra
popolazione neuronale che troviamo nelle corna anteriori ha un ruolo importante
nell'auto-controllo a livello spinale dell'eccitamento dei motoneuroni.
Vedremo poi che l'eccitamento dei neuroni è controllato anche a livello superiore nel controllo delle vie motorie ma vedete che c'è già un piccolo auto-controllo locale che ha come parte strutturale appunto le cellule di Renshaw.
Questi motoneuroni si trovano nelle corna anteriori del midollo a livello spinale ma non soltanto a livello spinale perché ricordiamo che noi adesso stiamo parlando delle corna anteriori del midollo spinale, costituite da motoneuroni che vanno ad innervare la muscolatura del tronco e degli arti. Dal punto di vista concettuale esattamente la stessa cosa sono i motoneuroni che troviamo nei nuclei motori dei nervi cranici, perché anche i muscoli della testa sono innervati in maniera motoria così come avranno le loro vie sensitive.
Ma non in tutti i nervi cranici, perché abbiamo detto c'è corrispondenza con i nervi spinali, ma tutto è un po' complicato nella morfologia della testa, ma in molti nervi cranici troviamo dei nuclei motori, i quali andranno ad innervare per esempio i muscoli mimici della faccia, i muscoli masticatori. Quei motoneuroni che troviamo nei nuclei motori dei nervi cranici sono esattamente equivalenti ai motoneuroni che troviamo nelle corna anteriori del midollo, solo che andranno ad innervare delle diverse popolazioni muscolari e non potranno essere innervate dal midollo perché stanno nella testa e quindi di loro si occuperanno i nervi cranici.
In ogni caso tutte questa popolazioni di motoneuroni vengono chiamate sia quelle che stanno nelle corna anteriori del midollo sia quelle che stanno nei nuclei motori dei nervi cranici, cioè in altre parole quelle che mandano gli assoni che vanno direttamente a raggiungere una cellula muscolare che determinano con il loro eccitamento, la contrazione della cellula muscolare, vengono chiamati secondo motoneurone oppure motoneurone inferiore perché nel controllo della motilità troveremo anche un primo motoneurone o motoneurone superiore che sarà disposto più in alto lungo il nevrasse e che andrà a controllare la funzione del secondo motoneurone. Accennavo prima alle vie motorie cioè quelle vie di controllo centrale della motilità per la postura e per il movimento volontario,le vie motorie saranno semplicemente costituite da assoni di motoneuroni che sono disposti più in alto rispetto a quelli di cui stiamo parlando, che saranno i primi motoneuroni.
VIA FINALE COMUNE
Quindi quando ci avviciniamo allo studio delle vie motorie dal punto di vista strutturale,ma che ha anche implicazioni a livello funzionale e clinico, come concetto generale dobbiamo pensare che sono presente dei secondi motoneuroni che sono quelli presenti nel corno anteriore del midollo o nel nucleo motore del nervo cranico, sono controllati e ricevono ordini da primi motoneuroni che sono a livelli superiori nel nevrasse.
Questo aspetto comporta un'altra definizione molto importante dal punto di vista funzionale e anche clinico tipico del secondo motoneurone: è quella di VIA FINALE COMUNE. I secondi motoneuroni costituiscono la via finale comune delle vie motorie nel senso che qualunque cosa accada all'interno del nevrasse,qualunque tipo di ordine,di regolazione che tutto il sistema nervoso centrale dalla corteccia cerebrale in giù con i circuiti più o meno complicati potrà dare, alla fine se il muscolo si contrae,questo si potrà contrarre soltanto se si eccita quel motoneurone, quella è la via finale comune di tutti i controlli motori dell'uno o dell'altro fascio muscolare.Questo significa che se quel motoneurone è morto o danneggiato, il muscolo non si potrà contrarre qualunque cosa accada in tutto il sistema nervoso centrale perché quella è la via finale comune, tutto arriva lì se vogliamo che il muscolo si contragga. Questo ha ovviamente delle implicazioni cliniche perché se il secondo motoneurone deve essere integro affinchè si contragga il muscolo,se c'è una lesione a carico del secondo motoneurone la sintomatologia non potrà che essere quella di una non contrazione del muscolo, di una paralisi flaccida.
Se la lesione invece non è a carico del secondo motoneurone che direttamente innerva il muscolo, ma a livello superiore delle vie di controllo non necessariamente l'esito sarà quello di una paralisi flaccida ma potrà essere e più spesso è, quella di una ipercontrazione del muscolo nel senso che in questo secondo caso quando la lesione è a carico delle vie superiori di controllo,cioè del primo motoneurone,allora il secondo motoneurone è in grado perfettamente di far contrarre il suo muscolo ma quello che viene a mancare è una corretta regolazione di questa contrazione. Questa corretta regolazione potrà esitare in una ipocontrazione o anche in una eccessiva contrazione e questo sarà il caso di una paralisi spastica. Quindi questa gerarchia delle due popolazioni neuronali, dei due tipi neuronali delle vie motorie avrà una diversa conseguenza nella clinica della lesione. Una lesione del secondo motoneurone sarà sicuramente una paralisi flaccida, una lesione del primo motoneurone potrà essere tra le più flaccida ma anche più spesso potrà essere spastica,che prevede un cattivo controllo non una mancanza radicale di funzione.
UNITA' MOTORIA
A questi due concetti molto importanti si accompagna un altro concetto che è nello stesso tempo strutturale e funzionale che è quello di unità motoria. Il concetto di unità motoria corrisponde ad un motoneurone e insieme con esso tutte le fibrocellule muscolari che questo motoneurone innerva,diciamo il territorio di competenza di quel neurone. Avremo delle unità motorie sia a livello dei nuclei che innervano il tronco sia a livello dei nuclei che innervano gli arti sia a livello dei nuclei che innervano le nostre diverse popolazioni muscolari della faccia. Nel caso dell'unità motoria la struttura,le dimensioni e la funzione sono legate in maniera molto diversa. All'unità motoria è legata la capacità di un certo distretto muscolare di svolgere movimenti fini oppure movimenti molto grossolani e ciò è direttamente legato alle dimensioni dell'unità motoria perché l'unità motoria,nel caso di una regione che compie movimenti molto fini, per esempio la mano, rispetto ad una regione che compie movimenti grossolani,per esempio il tronco,l'unità motoria dovrà essere più piccola. Immaginate cento fibre muscolari: se tutte queste sono innervate da un unico motoneurone, necessariamente dovranno essere o tutte contratte o tutte rilasciate. Se queste cento fibre muscolari sono innervate a dieci a dieci da dieci diversi motoneuroni,allora dieci fibre potranno essere contratte, dieci rilasciate e questo sarà un movimento fine. Quindi quanto più piccola è l'unità motoria, tanto più fine potrà essere il movimento di questa regione perché si potranno modulare contrazioni e rilasciamenti di popolazioni di fasci muscolari molto vicini tra di loro.
Se l'unità motoria è grande, tutti i fasci muscolari saranno contemporaneamente o tutti contratti o tutti rilasciati e allora necessariamente questa sarà una regione dove i movimenti sono grossolani. Nel tronco avremo delle grandi unità motorie,invece nelle regioni che innervano la mano avremo delle piccole unità motorie. Ciò significa che per innervare la mano occorre un numero di neuroni molto maggiore di quelli che occorreranno per l'innervazione del tronco. Quando esamineremo ad alti livelli le diverse somatotopie,vedremo che c'è un numero di neuroni molto maggiore che svolge la funzione di innervare la mano perché ognuna innerva poche fibre muscolari; mentre invece ci saranno pochi neuroni che dovranno innervare il tronco perché ogni neurone innerverà una grande quantità di fibre muscolari.
I RIFLESSI MONOSINAPTICI
La più semplice delle interazioni tra quello che accade a livello del corno posteriore e a livello del corno anteriore sono i cosiddetti riflessi monosinaptici .Del concetto di riflesso ne abbiamo già parlato: ad uno stimolo sensitivo corrisponde una risposta che può essere motoria o di altra natura. Il più semplice sarà proprio un riflesso monosinaptico che prevede un'interazione tra due cellule:una a livello del ganglio sensitivo e un'altra a livello del corno anteriore del midollo. Il più classico è il riflesso patellare, quello classico della visita neurologica in cui appunto si determinerà una distensione a cui fa seguito una risposta motoria.
Molti riflessi invece saranno mediati da numerose cellule nervose che si interpongono fra la stimolazione sensitiva e la risposta che può essere motoria, quindi in questo caso abbiamo semplicemente un interneurone che si interpone, ma potrà essere molto maggiore della quantità di cellule che si frappongono fino ad arrivare ai livelli superiori.
Esistono dei riflessi molto più complessi che esiteranno per la via sensitiva, per l'afferenza nervosa, una grande quantità di cellule nervose interposte a livello encefalico e una risposta che sarà di tipo neurosecernente.
|
Privacy |
Articolo informazione
Commentare questo articolo:Non sei registratoDevi essere registrato per commentare ISCRIVITI |
Copiare il codice nella pagina web del tuo sito. |
Copyright InfTub.com 2025