|
|
| |
ELEMENTI DI GENETICA
VARIABILITA' INDIVIDUALE: CARATTERI QUALITATIVI E QUANTITATIVI
La variabilità individuale è determinata sia da caratteri di tipo qualitativo che da caratteri di tipo quantitativo. I caratteri qualitativi sono, di solito, differenze di tipo alternativo che causano una variabilità di tipo discontinuo (il colore nero o rosso del mantello di certe razze di bovini, l'uniformità o la pezzatura del mantello, la presenza o l'assenza di corna, ecc.). I caratteri qualitativi, pur non avendo per lo più interesse pratico sono più facili da studiare, perché dipendono esclusivamente dal patrimonio ereditario non essendo influenzabili da fattori ambientali. I caratteri quantitativi sono invece differenze misurabili (la statura, il peso, la quantità di latte prodotta), sono più importanti da un punto di vista economico e sono fortemente influenzati dai fattori ambientali.
LEGGI DI MENDEL
Gregorio Mendel eseguì le sue osservazioni sui piselli odorosi, piante annuali ermafrodite che possono essere sia autofecondate che incrociate fra loro. Ne studiò i caratteri qualitativi (colore e forma dei semi, baccelli, colore dei fiori, ecc.). Selezionò delle linee pure, il colore dei semi si può presentare sotto due alternative: o giallo o verde, per ottenere una linea pura di piante a semi gialli, incrociò piante a semi gialli tra loro e tra i figli fece riprodurre solo quelli a semi gialli fino ad avere la certezza di avere ottenute piante pure per il carattere giallo; egualmente fece per il carattere colore verde dei semi.
Incrociò le due linee pure:
F1 = prima generazione filiale 100% semi gialli -carattere dominante-
F2 = seconda generazione filiale ¾ semi gialli e ¼ semi verdi -carattere recessivo-
Ogni carattere è determinato da un fattore ereditario (=gene) che può presentarsi sotto due forme differenti, dette alleli. I fattori ereditari si trasmettono da una generazione all'altra per mezzo dei gameti. L'individuo che ha due alleli uguali si dice omozigote, mentre eterozigote se ha due alleli diversi.
Le piante di partenza sono omozigote, ognuna per il suo carattere. Il fenotipo colore giallo del seme può essere determinato da un genotipo sia omozigote (GG) sia eterozigote (Gg), mentre il fenotipo colore verde dei semi è determinato solo dal genotipo omozigote gg.
La prima legge di Mendel, detta legge della dominanza, afferma che dei caratteri costituenti una coppia d'alleli l'uno, detto dominante, prevale sull'altro, detto recessivo.
La seconda legge, detta legge della segregazione o della purezza dei gameti, afferma che i due alleli si disgiungono alla formazione dei gameti, i quali quindi presentano uno solo degli alleli per ciascun carattere. Questa seconda legge è sempre valida. (Mitosi = dimezzamento dei cromosomi e quindi dei geni)
Le prime due leggi di Mendel si riferiscono a monoibridi, cioè ad incroci tra organismi diversi per una coppia d'alleli. Ci possono essere anche d'ibridi e poliibridi, cioè incroci interessanti due o più coppie d'alleli.
QUADRATO DI PUNNETT: G= giallo g= verde L= liscio l= rugoso
Gameti maschili
GL Gl gL gl
GL GGLL GGLl GgLL GgLl 100%
Gameti femminili
Gl GGLl GGll GgLl Ggll G = 75% L=50%
gL GgLL GgLl ggLL ggLl G = 75% L = 100%
gl GgLl Ggll ggLl ggll G = 50% L= 50%
La terza legge di Mendel, detta anche legge dell'indipendenza, afferma che negli incroci riguardanti più di una coppia d'alleli, ogni coppia d'alleli si comporta indipendentemente dalle altre, ricombinandosi nelle generazioni successive secondo le leggi della probabilità. Questa legge è valida solo se le coppie d'alleli sono portate da coppie diverse di cromosomi.
BASI CITOLOGICHE DELL'EREDITARIETA'
I geni sono posti nei cromosomi e gli alleli si trovano in coppia perché i cromosomi sono a coppie. Ogni individuo riceve un allele dal padre e l'altro dalla madre, la separazione d'ogni coppia di cromosomi omologhi è indipendente da quella delle altre coppie.
REINCROCIO
Il reincrocio è un test mediante il quale s'individua il genotipo sconosciuto di un fenotipo noto
? X omozigote recessivo
DOMINANZA INCOMPLETA O EREDITARIETA' INTERMEDIA
Non sempre la prima legge della dominanza appare valida. Per molte coppie d'alleli può non sussistere la dominanza di un allele rispetto all'altro e si manifesta un fenotipo intermedio (Es: mantello in una razza di bovini, mantello bianco + mantello rosso = mantello di colore roano, una colorazione intermedia fra il bianco e il rosso dei genitori). Dalla seconda generazione si può notare che non vi è mescolanza di fenotipo. La prima legge di Mendel viene quindi rinominata legge dell'uniformità degli ibridi di prima generazione.
EREDITA' INFLUENZATA DAL SESSO
In molti casi la dominanza o la recessività sono influenzati sia da fattori esterni (la temperatura) sia da fattori interni all'individuo (influenze ormonali). L'ereditarietà influenzata dal sesso si verifica quando l'espressione di un allele è condizionata dalla presenza nell'individuo di un ormone sessuale. La presenza di corna in alcune razze di pecore è influenzata dalla presenza di testosterone, negli omozigoti per la presenza di corna entrambi i sessi hanno questo carattere, mentre negli omozigoti per l'assenza di corna, le corna mancano in entrambi i sessi; negli ibridi eterozigoti l'allele presenza di corna domina sull'allele assenza di corna solo negli individui maschi in quanto evidentemente l'ormone maschile prodotto dal testicolo (testosterone) condiziona questa dominanza.
L'ereditarietà influenzata dal sesso o più precisamente la dominanza condizionata da ormoni sessuali è un'eccezione alla prima legge di Mendel, in quanto gli ibridi non sono uniformi.
Un altro esempio può essere anche la calvizie precoce nell'uomo, più che nella donna.
ALLELI LETALI
La presenza d'alleli allo stadio omozigot 959g69j e può risultare letale per l'organismo; la morte può avvenire molto precocemente, oppure può avvenire più tardivamente, ma sempre prima del raggiungimento della maturità sessuale e quindi prima della capacità procreativa. (Es: razza di topini a pelo giallo)
ALLELIA MULTIPLA
Un gene di solito si presenta sotto due forme all'eliche, ma in certi casi può avere più di due alleli; si parla allora di allelia multipla. Un esempio è dato dai principali gruppi sanguigni, infatti, abbiamo quattro fenotipi che determinati da un unico gene, che si presenta con tre alleli: La, Lb, Lo; vi è normale dominanza di La su Lo e di Lb su Lo, mentre tra La e Lb non vi è né dominanza, né recessività, né una condizione di dominanza incompleta, bensì entrambi gli alleli si esprimono.
PLEIOTROPIA
S'intende quel fenomeno genetico nel quale un solo gene influenza più caratteri diversi. Nel moscerino della frutta un allele che determina, allo stato recessivo, ali di dimensioni ridotte, influenza anche la forma dei bilancieri, la struttura degli organi genitali e la durata della vita del moscerino. Un altro esempio può essere l'albinismo nei topi.
EREDITA' DEI CARATTERI QUANTITATIVI: POLIMERIA O EREDITA' POLIGENICA
I caratteri più importanti da un punto di vista economico sono i caratteri quantitativi, cioè quelle caratteristiche dell'organismo che si possono misurare e che si manifestano secondo una variabilità continua.
Con il termine polimeria s'intendono quei casi di eredità nei quali un carattere quantitativo è determinato dalla somma dell'azione di più coppie di alleli, che si ereditano indipendentemente l'una dall'altra e fra i quali non vi è' dominanza e recessività.
Per esempio la lunghezza delle orecchie di un coniglio, incrociando una razza con orecchie molto lunghe ad una che le ha corte, nella prima generazione si ottiene una lunghezza intermedia, mentre nella seconda generazione si osserva una variabilità continua. Quindi s'ipotizza che il carattere sia determinato da tre coppie di alleli. Un altro esempio può essere il colore della pelle umana, anch'esso è determinato da tre coppie di alleli.
Altri caratteri quantitativi sono comandati da decine di geni e sono inoltre fortemente influenzati dall'ambiente.
DETERMINAZIONE DEL SESSO
Le differenze fra maschio e femmina sono causate da un corredo cromosomico diverso.
Eterogametia maschile di tipo XY
In molti gruppi animali le cellule somatiche e gli spermatogoni dei maschi presentano nel corredo cromosomico una coppia di cromosomi costituita da un grosso cromosoma X e da uno più piccolo Y; chiamati cromosomi sessuali o eterocromosomi e fanno sì che il maschio produca metà degli spermatozoi col cromosoma sessuale X e metà Y. Nella femmina la coppia di cromosomi sessuali è composta da due cromosomi X. I cromosomi apolidi eguali nei due sessi sono chiamati autonomi e indicati con la lettera A. La specie umana per esempio presenta tutte le cellule con 23 coppie di cromosomi delle quali 22 coppie sono autonomi, mentre la coppia di cromosomi sessuali è XY nel maschio e XX nella femmina, quindi AA + XY e AA + XX.
Il sesso del futuro bambino viene quindi stabilito all'atto della fecondazione.
Il sesso eterogametico è quello del maschio in quanto è il maschio che produce gameti diversi; questa determinazione è detta singamica poiché è all'atto della fecondazione che viene stabilito geneticamente il sesso dell'individuo.
Eterogonia maschile di tipo X0
E' un tipo di determinazione cromosomico del sesso che deriva da quello di tipo XY, solo che i maschi sono privi del cromosoma Y e hanno quindi un cromosoma sessuale in meno rispetto alle femmine; questa determinazione del sesso viene detta di tipo X0. I maschi sono AA + X e producono spermatozoi o A+ Y o A, le femmine sono AA + XX e fanno uova tutte A+ X.
Eterogametia femminile di tipo ZW
In questo tipo di determinazione del sesso le femmine hanno una coppia di cromosomi sessuali diversi tra loro (ZW), mentre il maschio ha i cromosomi sessuali uguali (ZZ); la femmina AA + ZW è eterogamica e produce uova A + Z o A + W, mentre il maschio omogametico AA + ZZ produce spermatozoi sempre A + Z.
Questa determinazione del sesso è progamica in quanto il sesso del futuro nascituro è già stabilito prima della fecondazione.
Tipo apolide-diploide
Un altro particolare tipo di determinazione cromosomica del sesso è di tipo apolide-diploide che è presente nella maggioranza degli Imenotteri (api). Le femmine sono diploidi, mentre i maschi sono aploidi. Le uova prodotte dalla femmina possono svilupparsi sia se fecondate sia se non fecondate: nel primo caso si svilupperanno delle femmine, nel secondo caso si svilupperanno per partenogenesi dei maschi, i quali essendo apolidi, avranno una spermatogenesi senza meiosi.
NATURA QUANTITATIVA DELLA DETERMINAZIONE CROMOSOMICA DEL SESSO
Si cercò di stabilire quanti geni fossero implicati nella determinazione del sesso e su quali cromosomi questi geni fossero collocati.
Dal moscerino della frutta si osservò che i maschi erano AA + XY e le femmine AA + XX, ma si riscontravano talvolta degli individui con corredo cromosomico anormale che presentavano diverse anomalie nella sessualità. Triplodia degli autonomi e un unico cromosoma sessuale X, AAA + X sono maschi sterili che presentano l'apparato genitale molto sviluppato. I geni che comandano la sessualità quindi non sono sul cromosoma Y, questi geni di mascolinità sono collocati sugli autonomi, poiché la triplicazione di questi aumenta l'espressione dei caratteri maschili. Altri individui anormali avevano triplodia del cromosoma sessuale X, AA + XXX, sono femmine sterili che presentano l'apparato riproduttore molto sviluppato, quindi i geni della femminilità sono posti sul cromosoma X, dato che con una sua triplicazione aumenta l'espressione dei caratteri femminili.
In un maschio normale del moscerino AA + XY i geni di mascolinità posti sugli autonomi sono più numerosi o sono più forti dei geni della femminilità posti su un unico cromosoma X, e viceversa, quelli di una femmina normale AA + XX sono più forti di quelli di un maschio anormale.
Esistono infine individui anormali con triplodia degli autonomi e normale coppia di cromosomi sessuali X. Questi individui AAA + XX sono sterili e con caratteri sessuali intermedi ai due sessi. In questi intersessi i geni della mascolinità sono in ugual numero hanno forza uguale a quelli della femminilità.
L'essere maschio o l'essere femmina è quindi una questione di carattere quantitativo, dato che ogni maschio porta anche una minoranza di geni di femminilità e ogni femmina una minoranza di geni di mascolinità. Questa situazione di tipo quantitativo è valida per quasi tutti gli animali a sessi separati.
Nell'uomo e in altri mammiferi i geni della femminilità sono portati dai cromosomi sessuali X, mentre i geni della mascolinità sono portati in massima parte dal cromosoma sessuale Y.
Nella specie umana ci sono talvolta individui con situazioni patologiche dovute a corredo cromosomico anormale di tipo AA + XXY; questi individui sono maschi sterili con testicoli poco sviluppati; possono presentarsi anche individui di tipo AA + X che sono femmine sterili con ovari iposviluppati.
Negli animali a sessi separati vi è prevalenza dei geni di un sesso rispetto all'altro, mentre negli animali ermafroditi i geni di mascolinità e quelli di femminilità sono in equilibrio.
Vi sono però anche delle specie animali che non hanno una determinazione del sesso genotipicamente stabile e nelle quali il sesso viene determinato dopo la fecondazione da fattori esterni come la temperatura o l'alimentazione, questo tipo di determinazione del sesso viene detto metagamico. Questo spiega anche come sia possibile un'inversione di sesso spontanea. Non bisogna confondere queste inversioni sessuali spontanee e fisiologiche da quelle provocate da malattie.
Un altro esempio di inversione sessuale causata da influenze ormonali esterne è dato dai così detti free-martins, che sono bovini sterili. Quando tra due feti gemelli di bovini di sessi geneticamente diversi si stabiliscono delle connessioni vascolari tra le placente, il feto di sesso femminile, a causa dell'ormone maschile che riceve dal gemello, inverte il sesso e diventa un maschio che però è sterile.
EREDITA' LEGATA AL SESSO
Si tratta di quei caratteri determinanti da geni situati sui cromosomi sessuali e che si trasmettono in modo differente a seconda se provengono dal padre o dalla madre. Un esempio di eredità legata al sesso è come si eredita nell'uomo il daltonismo; il carattere per il daltonismo e quello per la visione normale sono comandati da un gene posto sul cromosoma sessuale X del maschio. Risulta chiaro che il daltonismo è più frequente nel sesso maschile dato che femmine daltoniche possono nascere solo da matrimoni nei quali o entrambi i genitori sono daltonici oppure il padre è daltonico e la madre portatrice sana.
ASSOCIAZIONE E SCAMBIO
Tutti i geni che si trovano su uno stesso cromosoma si dicono associati e vengono ereditati insieme; questo fenomeno, denominato associazione genica, rappresenta una significativa eccezione alla terza legge di Mendel, poiché si possono ereditare in modo indipendente solo quei geni che sono posti su cromosomi diversi. Il crossing over, cioè lo scambio di alleli tra cromosomi omologhi avviene durante la profase della prima divisione meiotica.
L'associazione genica provoca il fatto che i geni si ereditino in prevalenza a gruppi, mentre il crossing over o scambio rompe quest'associazione e ripristina parzialmente l'indipendenza delle coppie d'alleli, con il vantaggio evolutivo di creare una maggiore variabilità.
MUTAZIONI
Il patrimonio ereditario di un organismo è generalmente molto stabile, la molecola di DNA tende sempre a duplicarsi in modo conservativo. Talvolta però può accadere che il DNA duplicandosi non faccia una copia esatta di se stesso o che un singolo cromosoma cambi forma o che si modifichi il numero cromosomico; in tutti questi casi si parla di mutazioni, cambiamenti del patrimonio ereditario. Improvvisi cambiamenti del patrimonio ereditario che possono essere trasmessi alla discendenza solo se interessano le cellule germinali.
Esistono tre principali mutazioni: gnomiche, cromosomiche e geniche.
Mutazioni gnomiche
Sono cambiamenti nel numero dei cromosomi di una determinata specie e causano quindi imponenti variazioni nel numero dei geni. Queste mutazioni possono consistere nella presenza tripla di un singolo cromosoma oppure nella perdita di un cromosoma.
Mutazioni cromosomiche
Sono cambiamenti nella struttura di un cromosoma che causano quindi variazione nel numero e nell'ordinamento dei geni posti in quel cromosoma. Spesso producono sterilità.
Mutazioni geniche
Sono cambiamenti nella struttura chimica del gene e consistono in variazioni nella sequenza delle basi azotate della molecola di DNA. Si possono considerare come errori di copiatura durante la duplicazione del DNA.
Le mutazioni sono uno dei mezzi più importanti per lo studio della genetica in quanto la conoscenza della struttura e funzione dei geni è possibile solo col confronto tra le forme alternative dei geni.
LA TEORIA DELL'EVOLUZIONE
La teoria dell'evoluzione, una delle scoperte
scientifiche che hanno influito più profondamente sulla cultura moderna e sulla
concezione del mondo dell'uomo contemporaneo, fu concepita e messa a punto,
nelle sue linee essenziali, da Charles Darwin nel corso
dell'Ottocento, in un periodo di grandi progressi nelle scienze della natura.
Nel XVIII secolo diversi scienziati e filosofi avevano cominciato a mettere in
discussione la concezione di un mondo immutabile: ad esempio Kant nel 1775
aveva formulato l'ipotesi secondo cui il sistema solare trarrebbe origine dal
moto vorticoso di una nebulosa primitiva; quest'ipotesi sarà ripresa alla fine
del secolo da Pierre Simon de Laplace.
Nella seconda metà del Settecento viaggi, spedizioni scientifiche sistematiche
ed esplorazioni, seppur motivate principalmente da scopi commerciali, avevano
dato un forte impulso alla ricerca in campo biologico e fatto nascere la
paleontologia e la geologia
che, con gli studi di Charles Lyell e Georges Cuvier, avevano rivelato strati
geologici formatisi in tempi successivi, che incorporavano i resti di specie
animali e vegetali ormai scomparse da tempo dalla Terra. In biologia, lo
sviluppo dei metodi di analisi comparata degli organismi rendeva sempre più
inverosimile concepire le specie viventi come fisse e immutabili, mentre la
scoperta di animali difficilmente classificabili negli schemi esistenti
imponeva con sempre maggior forza l'idea dell'evoluzione delle specie.
L'evoluzionismo scientifico nacque con il francese Jean-Baptiste de Lamarck
(1809: Filosofia zoologica), secondo il quale i caratteri acquisiti durante la
vita dell'individuo possono essere trasmessi ai discendenti (ereditarietà dei
caratteri acquisiti); oggi questa teoria è stata abbandonata, perché non ha
trovato verifiche sperimentali convincenti.
Di fondamentale importanza fu l'opera di Charles Darwin
(1809-1882) che dedicò tutta la vita a raccogliere materiale di studio, per
giungere alla formulazione di una teoria dell'evoluzione che avesse un solido
fondamento scientifico. Nel 1831 s'imbarcò come naturalista di bordo sul
brigantino Beagle; il viaggio intorno al mondo durò quasi cinque anni, durante
i quali Darwin raccolse un'ingente quantità di campioni ed eseguì numerose
osservazioni, che costituirono la base per elaborare la teoria dell'evoluzione
delle specie, contrapposta alla concezione "fissista".
Contributi altrettanto importanti furono dati da Darwin con la messa a punto
dei concetti di evoluzione ramificata, che implica la discendenza da un'origine
comune di tutte le specie viventi, e di evoluzione graduale, contrapposta a
quella a salti (mutazionismo) che sosteneva come una nuova specie si afferma
attraverso una singola mutazione. Netta era l'esclusione di qualunque
interpretazione di tipo finalistico, sostenuta da altre teorie evolutive (ad
esempio quella di Lamarck): nella sua opera principale, On the Origin of
Species by Means of Natural Selection, utilizzò sempre l'espressione
"discendenza con modificazione", probabilmente perché riteneva che la
parola "evoluzione" suggerisse un legame troppo stretto con il
concetto di progresso.
La svolta teorica fu ancora più decisiva con
l'affermazione del principio della selezione naturale, che restò a lungo per
molti "soltanto un'ipotesi" anche quando la teoria dell'evoluzione
era già stata largamente accettata. Secondo Darwin, il meccanismo della
discendenza con modificazioni avviene in due fasi: dapprima si ha lo sviluppo
di un'abbondante varietà di individui, che vengono poi selezionati tramite il
criterio della sopravvivenza del più adatto, o selezione naturale. La prima
fase è dominata dalla casualità, la seconda dalla necessità.
La controversia fra le diverse teorie evolutive durò diversi decenni; le
ricerche nel campo della genetica (genetica evoluzionista) e, in seguito, della
biologia molecolare, fornirono poi molti argomenti a favore del darwinismo. Un
notevole contributo venne anche dallo sviluppo di nuovi criteri tassonomici (v.
riquadro) e dalla paleontologia.
Ormai il darwinismo è accettato quasi unanimemente, almeno in ambito
scientifico; il dibattito, a volte anche aspro, verte principalmente sul peso
da attribuire ai diversi fattori che agiscono sull'evoluzione.
Nel rapporto fra casualità e necessità, ad esempio, Richard Dawkins e Daniel
Dennett privilegiano la seconda, nella convinzione che la selezione naturale
regoli tutto ciò che ha qualche importanza nell'evoluzione, riducendo di molto
il ruolo ricoperto dalla fase della variazione. Dawkins ha costruito una teoria
generale della trasmissione dei tratti culturali, che avverrebbe tramite i
"memi", presunti equivalenti dei geni, che si replicano utilizzando
le menti umane. In questa concezione è vicino a Edward O. Wilson, padre della
sociobiologia, che costituisce un tentativo di spiegare biologicamente la
cultura e l'organizzazione sociale.
L'impostazione concettuale che privilegia la fase della necessità viene
definita fondamentalismo darwiniano da Stephen Jay Gould e Niles Eldredge che,
accentuando invece l'aspetto della casualità, sostengono la teoria degli
"equilibri punteggiati": secondo questa teoria l'evoluzione non
procede con un cambiamento lento e costante, ma con l'alternanza di lunghi
periodi di stasi e repentini cambiamenti, a volte dovuti a eventi catastrofici,
come la scomparsa dei dinosauri che sarebbe stata causata dall'impatto di un
gigantesco meteorite.
Il genetista Richard C. Lewontin è molto vicino a
Gould nel criticare le posizioni del determinismo genetico e della
sociobiologia, e propone una visione dialettica del rapporto fra le parti e il
tutto che, tenendo ben salde le conquiste del pensiero scientifico, superi i
limiti della concezione riduzionista, consentendoci di vedere tutta la
ricchezza delle interazioni che si presentano in natura.
Tutto questo rende la teoria dell'evoluzione un complesso di conoscenze vasto e
articolato, che ha fecondato la ricerca in numerosi rami della scienza
naturale, ricevendone conferme, occasioni di nuovi sviluppi e correzioni. Anche
per il futuro l'evoluzionismo potrà rappresentare il filo conduttore in grado di
garantire l'interpretazione dell'immensa quantità di dati raccolti dalla
ricerca genetica e in altri rami della scienza.
Il biologo francese François Jacob, premio Nobel 1965, ha paragonato l'evoluzione all'opera di chi si diverte a fare "bricolage", accumulando tutto ciò che trova per creare nuovi prodotti. Attraverso una serie di meccanismi genetici e ambientali, alcuni pezzi vengono adoperati per fare cose diverse, o vengono modificati a seconda delle varie esigenze. E' curioso osservare, per esempio, come nei vertebrati gli arti anteriori si siano modificati per rispondere, partendo da un modello comune, a diverse esigenze funzionali: attraverso gli stessi elementi di base (le ossa degli arti), l'evoluzione ha modellato, attraverso graduali modificazioni, strumenti molto diversi: pinne, ali, zampe e braccia. Dalle pinne pettorali di un comune antenato, l'Eusthenopteron (il pesce che per primo "sbarcò sulla terra" ed è un progenitore degli anfibi), si pensa infatti che derivino le ali del gabbiano, le pinne pettorali del delfino, le zampe anteriori del cane e le braccia dell'uomo. L'ipotesi di un continuo divenire e trasformarsi del mondo e degli esseri viventi, oggi comunemente accettata, venne formulata scientificamente nel secolo scorso, e suscitò molte polemiche. Fino allora si pensava, secondo un'interpretazione letterale della Genesi, che Dio avesse creato ogni essere così com'era e come sarebbe sempre stato. Il grande naturalista svedese Carlo Linneo (1707-1778), all'inizio della sua grande opera di classificazione biologica, poteva ancora affermare: "Numeriamo tante specie quante in principio furono create dall'Ente infinito". La svolta avvenne con la pubblicazione de L'origine delle specie di Charles Darwin (1859), che segnò un punto di non ritorno: la teoria dell'evoluzione sarebbe divenuta, con opportune variazioni e grazie al fecondo incontro con la genetica all'inizio del nostro secolo, una delle teorie portanti della scienza contemporanea. Il collo della discordia La prima teoria organica dell'evoluzione fu sostenuta da Jean Baptiste de Lamarck (1744-1829), secondo il quale le varie specie animali sono soggette a trasformazioni a causa dell'azione dell'ambiente, che determina la comparsa di caratteri ereditari tramite il principio dell'uso o non uso di un organo: l'uso farebbe sviluppare l'organo, mentre il non uso lo indebolirebbe fino a farlo scomparire. Il lungo collo della giraffa, per esempio, sarebbe dovuto al fatto che quell'animale vive in un ambiente povero di erba, che lo obbliga a brucare il fogliame degli alberi, sforzandosi di arrivare in alto. "Da quest'abitudine, mantenutasi per lungo tempo in tutti gli individui della sua razza, è risultato che le zampe anteriori sono diventate più lunghe di quelle posteriori, e che il suo collo si è tanto allungato da permetterle di alzare la testa toccando i sei metri di altezza senza drizzarsi sulle zampe posteriori". Nei vari organismi, inoltre, esisterebbe un impulso interno, quasi una volontà, di trasformarsi in forme più evolute.
La teoria lamarckiana venne contestata dal famoso e autorevole naturalista Georges Cuvier (1769-1832), che credeva nella fissità delle specie, mentre l'esempio della giraffa venne deriso da una serie di vignette apparse sul giornale francese Figaro: Lamarck avrebbe passato i suoi ultimi anni dimenticato, in solitudine e povertà. Eppure il suo modello, anche se diverso da quello che si affermò poi con Darwin, offriva una spiegazione molto semplice dei fenomeni di adattamento all'ambiente e dell'evoluzione: forse per questo è stato riproposto, sia pure con modifiche, a varie riprese. A volte in modo drammatico, come nel caso del biologo austriaco Paul Kammerer (1880-1926), che condusse degli esperimenti su diverse generazioni di "rospo ostetrico" per dimostrare l'"ereditarietà dei caratteri acquisiti", e sostenne di averne trovato la prova, innescando una vivace polemica con altri scienziati; quando si scoprì che l'ultimo esemplare rimasto in suo possesso era stato contraffatto, si uccise. All'inizio de L'origine delle specie, Darwin sottolineò i meriti di Lamarck; tuttavia "né l'azione dell'ambiente né la volontà degli organismi" gli sembrarono mai sufficienti a spiegare la complessità dell'evoluzione. Oggi sappiamo che, peraltro, i caratteri acquisiti "per abitudine" durante la vita non possono modificare il patrimonio genetico. Il punto di partenza della teoria di Lamarck è l'estrema variabilità degli organismi soggetti a domesticazione o viventi allo stato naturale. Le variazioni tra individuo e individuo di una stessa specie (di cui Darwin ignorava l'origine) risultano determinanti per l'evoluzione futura perché individui e specie sono immersi in un ambiente in cui c'è una cronica e crescente povertà di risorse. Quest'ultimo principio, espresso dall'economista Thomas Malthus (1766-1834) nei riguardi della popolazione umana, suggerì a Darwin il concetto di "lotta per l'esistenza": individui e specie si fanno spazio a spese di altri, e gli individui vincitori di questa lotta, che riescono non solo a sopravvivere ma anche a trasmettere i propri caratteri alle generazioni successive, sono gli individui più adatti o meglio adattati alle condizioni ambientali. Questo meccanismo, che costituisce la "selezione naturale", esclude ogni intervento di impulsi o volontà, ed evita l'idea di un'azione diretta dell'ambiente: si tratta di un fenomeno scientificamente osservabile e in parte misurabile. La teoria di Darwin, pur tra fiere resistenze, incontrò un sempre maggiore successo, fino a diventare dominante. Con lo sviluppo della genetica, a partire dall'inizio del nostro secolo, si è arrivati all'idea di un processo evolutivo che avviene gradualmente da specie antenate (o "ancestrali") attraverso la combinazione della selezione naturale e delle mutazioni del patrimonio genetico. Il ritmo temporale uniforme, lento e graduale (secondo il principio di continuità per cui "la Natura non fa salti"), tuttavia, è stato messo in discussione dalla cosiddetta teoria degli "equilibri punteggiati", secondo cui le nuove specie si sviluppano in modo improvviso e rapido, dopo lunghi periodi di stasi. Il processo evolutivo dell'okapi e della giraffa a partire da un antenato comune (pre-okapi), per esempio, può essere rappresentato da un modello "gradualista" o da uno in cui il mutamento è improvviso, e si accompagna alla formazione di nuove specie isolate. A 22 anni Charles Darwin ottenne un posto di naturalista (non pagato) a bordo del brigantino Beagle (ossia "bracchetto": oggi si chiamerebbe Snoopy), che per ordine dell'Ammiragliato inglese doveva compiere una spedizione scientifica intorno al mondo.
Il viaggio durò cinque anni e, come scrisse lo stesso Darwin, fu certamente l'avvenimento più importante della sua vita, che decise tutta la sua futura carriera. Il 15 settembre 1835 il Beagle arrivò in vista delle isole Galápagos, che devono il nome alle grandi tartarughe che vi abitano (in spagnolo appunto "galápagos"). Queste isole, scoperte casualmente nel 1535 dagli spagnoli, che le chiamarono anche Islas Encantadas, rimasero disabitate per molto tempo, ricevendo sporadiche visite di bucanieri e di cacciatori di balene, finché nel 1832 vennero annesse dalla repubblica dell'Ecuador, di cui fanno parte tuttora. Situate a circa 1000 km dalla costa sudamericana, si trovano sulla linea dell'equatore ma sono lambite dalla corrente fredda di Humboldt, che ne mitiga notevolmente la temperatura: per questo vi si è sviluppato un ambiente naturale particolare, unico al mondo. Il Beagle incrociò nelle acque dell'arcipelago fino al 20 ottobre, per poi fare rotta verso Tahiti: in questo periodo vennero effettuati degli sbarchi, in ordine di successione, sulle isole San Cristóbal (o Chatham), S. Maria (o Floreana o Charles), Isabela (o Albemarle) e San Salvador (o Santiago o James). Con la consueta diligenza, Darwin compì numerose osservazioni su tartarughe, lucertole e uccelli, rimanendo soprattutto colpito dai fringuelli, che presentavano becchi diversi a seconda delle isole in cui vivevano. I "fringuelli di Darwin", come in seguito furono chiamati, sono 14 specie che si ritengono evolute da un comune antenato, risalente a un'epoca compresa tra uno e cinque milioni di anni fa. Hanno piumaggio in prevalenza scuro e proporzioni simili, ma variano in lunghezza (da sette a 12 cm) e nella forma del becco, che riflette le diverse abitudini alimentari: come variano le pinze a seconda degli usi cui sono destinate, così variano i becchi a seconda che i fringuelli si nutrano di semi, di insetti o di fiori e frutti di cactus. Nel suo scrupoloso diario, Darwin scriveva che "vedendo questa graduazione e diversità di struttura in un piccolo gruppo di uccelli, strettamente affini, si potrebbe realmente immaginare che da un originario piccolo numero di uccelli di questo arcipelago sia stata presa una specie e modificata per diversi fini": la teoria dell'evoluzione incominciava a prendere forma. La teoria della selezione naturale presupponeva l'esistenza di un'estrema variabilità che veniva trasmessa ereditariamente: tuttavia ai tempi di Darwin, come egli stesso aveva lamentato, nulla si sapeva sulle leggi dell'ereditarietà e su quelle della variabilità. La teoria dell'ereditarietà oggi universalmente accettata venne proposta per la prima volta dal monaco Johann Gregor Mendel nel 1865, ma venne "riscoperta" e divenne nota nel 1900. Anche la comprensione del meccanismo attraverso cui si genera una nuova variazione ereditaria era destinata ad avere inizio nel nostro secolo, e solo nella seconda metà venne provato che il Dna è responsabile dell'informazione genetica degli organismi viventi e se ne scoprì la struttura, il modo in cui essa può cambiare e come da essa dipenda la sintesi delle proteine e lo sviluppo dell'intero organismo. Ogni tratto di Dna che "codifica" (cioè contiene l'informazione per sintetizzare) una determinata proteina costituisce un gene, il cui ruolo è quello di specificare la natura e l'ordine degli aminoacidi che la compongono.
La regione centrale di un gene (regione codificante) è costituita da una sequenza in cui si susseguono quattro componenti elementari (basi o nucleotidi), che vengono "letti" a gruppi di tre e "tradotti" nel linguaggio dei 20 aminoacidi che sono i componenti elementari delle proteine degli esseri viventi: ogni tripletta corrisponde a un aminoacido. Se nella regione codificante di un gene un nucleotide cambia natura, si modifica anche la relativa tripletta, e quindi viene codificato un aminoacido diverso, e la proteina risulta in genere alterata. Perché avviene la mutazione di un gene, che nel suo nuovo assetto passerà di generazione in generazione? Nella maggior parte dei casi senza un motivo specifico, semplicemente per un errore avvenuto al momento della replicazione del Dna. Il lungo filamento del Dna infatti si replica ogni volta che una cellula si deve dividere, in modo da trasmettere alle due cellule figlie la stessa informazione genetica della cellula madre: a volte accade un errore, e al posto di un nucleotide ne viene inserito un altro, che da quel momento resterà in quella posizione ogni volta che il Dna si replicherà. La frequenza degli errori è comunque bassa: in media viene inserito un nucleotide sbagliato su un miliardo. Le mutazioni quindi si verificano per lo più spontaneamente, con un ritmo in genere costante, misurabile nelle varie specie: nell'uomo ci sono due o tre mutazioni spontanee in ognuna delle nostre cellule sessuali. Il tasso di mutazione può essere più elevato in presenza di particolari agenti fisici o chimici (basti pensare ai casi di Seveso o Chernobyl). Quali conseguenze possono avere le mutazioni? In alcuni casi predispongono altre mutazioni, in altri possono far insorgere malattie, in altri ancora possono risultare vantaggiose. In termini evolutivi, è grazie alle mutazioni che le varie forme di vita si sono diversificate e trasformate, adattandosi agli ambienti più diversi. La selezione ha fatto sì che venissero eliminate numerose varianti inadatte e si affermassero quelle più idonee: attraverso questo meccanismo l'ambiente ha indirettamente modellato le caratteristiche di volta in volta necessarie agli esseri viventi. Le condizioni ambientali, infatti, sono cambiate più volte nei tempi lunghissimi dell'evoluzione, che ha via via "pescato" tra un immenso numero di combinazioni. Prendiamo in considerazione l'elefante, tipico animale equatoriale: nella sua discendenza ci sono varianti "nordiche", che vengono corrette dall'ambiente eliminando quelle troppo accentuate. In passato si può pensare che tali varianti siano state selezionate quando l'ambiente lo richiedeva: ecco così il mammut, sviluppatosi anche in climi molto freddi. Cambiando le condizioni ambientali, si sono affermati i portatori delle caratteristiche che meglio a esse si adattavano.
Secondo la teoria dell'evoluzione, gli esseri viventi hanno un'origine comune e si sono progressivamente trasformati e sviluppati attraverso un processo che dura ancora oggi. Al di là delle osservazioni dei naturalisti, vi sono delle "prove" esibite dalla biologia molecolare. Una di esse consiste nello studio di una particolare proteina, il citocromo c, di cui tutti gli organismi viventi hanno bisogno per ossidare le molecole dei cibi, con la respirazione, ottenendo energia.
La sua comparsa nel corso dell'evoluzione contribuì all'origine degli organismi multicellulari La molecola di citocromo c è costituita da 104 aminoacidi che circondano un gruppo "eme" a forma di rosetta, con un atomo di ferro al centro. La collocazione degli aminoacidi può variare (ferma restando la struttura tridimensionale della molecola): è identica nell'uomo e nello scimpanzé, mentre cambia (in 12 dei 104 siti) nell'uomo e nel cavallo, per raggiungere il massimo di differenza nell'uomo e nella muffa del pane (44 siti su 104). Come possono due catene proteiche così diverse avere funzioni simili? La risposta è che la maggioranza degli aminoacidi sono intercambiabili, mentre solo alcuni (35 in tutte le specie conosciute) non possono essere sostituiti senza che la proteina cessi la sua attività. Le differenze, d'altra parte, sono preziose dal punto di vista evolutivo: quanto più indietro nel tempo gli antenati di due organismi attuali si sono separati, tanti più cambiamenti si sono accumulati nella sequenza degli aminoacidi. Per la sua antichità e la facilità con cui può essere isolato, il citocromo c è stato oggetto di grande attenzione da parte dei biologi interessati all'evoluzione, così che possediamo moltissime informazioni sul suo sviluppo. E' stato così possibile tracciare degli alberi genealogici, che sono in accordo con quelli elaborati con metodi tradizionali: il confronto delle sequenze degli aminoacidi costituisce dunque un prezioso strumento per studiare i processi evolutivi.
Nel meccanismo della selezione naturale un ruolo non secondario è stato giocato dall'abilità di alcuni nel trovare nuove fonti alimentari, traendo energia da sostanze per altri indigeste o poco nutrienti. Nella natura quale oggi si presenta, le piante verdi sono le sole in grado di produrre materiali biologici utilizzando sostanze inorganiche; gli altri esseri viventi possono unicamente consumare materia organica. I vegetali costituiscono l'anello di partenza delle catene alimentari, che sono delle sequenze coordinate di organismi, nelle quali gli uni mangiano gli altri prima di essere a loro volta mangiati. Esistono più tipi di catene alimentari. Quelle dei predatori vanno dai vegetali agli erbivori, ai piccoli carnivori, ai carnivori più grossi e così via; quelle dei parassiti vanno dagli organismi di grandi dimensioni a quelli sempre più piccoli; quelle dei detritivori vanno dalla materia organica morta (detrito) agli organismi. In realtà non ci sono catene alimentari isolate, ma un complesso gruppo di connessioni alimentari che si intersecano e formano una rete alimentare, in cui ogni organismo, e in senso più ampio ogni specie, occupa una precisa posizione. Gli ecosistemi con uno scarso numero di specie presentano una rete alimentare semplice e molto sensibile ai cambiamenti, mentre quelli con numerose specie ne hanno una più equilibrata e stabile.
L'entomologo britannico C. B. Williams ha calcolato che in ogni istante sulla Terra il numero di insetti vivi è di circa un miliardo di miliardi. Due bravi e simpatici mirmecologi (studiosi delle formiche: al mondo non sono più di 500) hanno ragionato su questo dato traendo alcune conclusioni sulle creature cui hanno dedicato la loro vita di scienziati: se, come suggerisce una stima prudente, le formiche costituiscono l'uno per cento degli insetti, la loro popolazione complessiva è di 10 milioni di miliardi.
Poiché il loro peso varia, a seconda delle specie, tra uno e cinque milligrammi, messe tutte insieme peserebbero quanto tutti gli esseri umani, cui contendono il ruolo di organismi predominanti sulla terraferma: si tratta di una prospettiva particolare, ottenuta abbassando il punto di vista a un millimetro dalla superficie del suolo, e che forse ha influenzato la recente produzione di qualche film. Originatesi all'epoca dei dinosauri, le formiche hanno avuto una spettacolare diffusione di forme adattative: ne sono state classificate 9500 specie, dalla Finlandia all'Amazzonia, ma si pensa che probabilmente siano alcune decine di migliaia. Il calcolo delle specie non costituisce una scienza esatta: secondo il primo rapporto globale dell'Onu sulla biodiversità (pubblicato nel novembre 1995), le specie finora note e ufficialmente classificate sono 1.750.000, ma si stima che quelle esistenti possano essere più di 15 milioni. Tenendo conto che alcuni habitat, dove la varietà delle specie animali e vegetali è vastissima (come le volte delle foreste tropicali e i fondali oceanici), sono in buona parte ancora sconosciuti, c'è anche chi ipotizza che le specie esistenti possano essere più di 30 milioni o arrivare addirittura a 100 milioni. La biodiversità, e cioè la diversità delle forme viventi sulla Terra, è un elemento costitutivo dell'ambiente, nell'ambito del quale contribuisce alla stabilità del clima, alla regolazione del deflusso delle acque e a rinnovare la fertilità del suolo. Di qui l'allarme per quanto avviene in varie zone del nostro pianeta: molte specie sono scomparse o stanno scomparendo per i mutamenti indotti nel loro habitat. Nella sola foresta tropicale, per esempio, i biologi stimano che ogni ora si estinguano tre specie di esseri viventi. L'estinzione delle specie non è un fenomeno nuovo: si calcola infatti che il 99 per cento delle specie vissute sulla Terra si sia estinto, attraverso l'inesorabile meccanismo della selezione o in virtù di eventi catastrofici. L'ultima estinzione di massa avvenne circa 65 milioni di anni fa, e spazzò via i dinosauri e i due terzi di tutti gli animali esistenti. Il recupero richiese milioni di anni, ma portò a una proliferazione di specie maggiore che in ogni altra epoca. Secondo i biologi, oggi ci troviamo probabilmente alle soglie di un'altra estinzione di massa, provocata dall'uomo e di cui l'uomo subirà le conseguenze. Che possono essere anche economiche, dal momento che è dannoso eliminare possibilità che in futuro potrebbero rivelarsi vantaggiose (una specie selvatica di mais, "salvata" recentemente da un ricercatore, si è per esempio rivelata un affare da miliardi di dollari). Insomma: la biodiversità è necessaria e utile, al di là di certi ricorrenti e "toccanti" appelli per l'estinzione di alcune specie. Per questo oltre 150 Paesi hanno firmato nel 1992 una Convenzione internazionale per la salvaguardia e l'uso "sostenibile" della biodiversità: i risultati di questo accordo, a dire il vero, non sembrano particolarmente brillanti. A tutt'oggi almeno 5400 specie animali e vegetali rischiano di estinguersi.
PROVE DELL'EVOLUZIONE
Le prove scientifiche dell'evoluzione sono innumerevoli e comprendono prove ricavabili dallo studio dei fossili, prove deducibili dall'osservazione delle strutture degli organismi viventi, prove embriologiche, prove biogeografiche e fenomeni evolutivi direttamente osservabili.
Testimonianze fossili
La paleontologia ha dimostrato che non tutte le specie attualmente esistenti sulla Terra erano presenti anche in un lontano passato e che, per contro, molte specie un tempo esistenti sono ormai scomparse. I paleontologi sono in grado di stabilire l'età dei fossili e quindi di ricostruire la successione cronologica ed evolutiva delle specie i cui resti sono presenti nei fossili.
Per alcuni gruppi di animali i reperti fossili sono così abbondanti e ben
distribuiti nel tempo, da formare delle serie complete che consentono di
ricostruire in modo particolareggiato le tappe del loro processo evolutivo.

Per quanto riguarda i Vertebrati lo studio dei fossili testimonia che i primi vertebrati a comparire sono stati i Pesci seguiti dagli Anfibi, dai Rettili e infine dagli Uccelli e dai Mammiferi. Un eccezionale fossile che testimonia l'evoluzione degli Uccelli a partire dai Rettili è l'Archaeopteryx, nel quale caratteri rettiliani sono mescolati con quelli degli Uccelli. La derivazione degli Anfibi dai Pesci è testimoniata da un altro fossile, Ichtyostega che presenta caratteristiche intermedie tra una classe e l'altra.

Per quanto riguarda il mondo vegetale, la derivazione delle piante terrestri
dalle alghe pluricellulari marine è testimoniata dal ritrovamento di piante
fossili le cui caratteristiche sono intermedie tra le une e le altre.
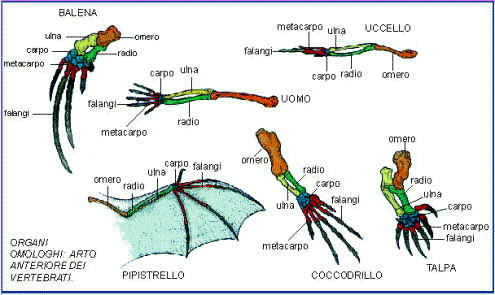
Omologia
Se si osserva l'arto anteriore di pesci, degli anfibi, dei rettili, degli uccelli e dei mammiferi, si può notare come questo sia in tutti i casi citati costituito dagli stessi pezzi scheletrici più o meno sviluppati: omero, radio, ulna, ossa carpali, ossa metacarpali e falangi. Dato il diverso utilizzo che le specie ne fanno, l'arto si è modificato in una moltitudine di forme diverse, ma in queste diverse classi di vertebrati è sempre riscontrabile la stessa struttura. Questo tipo di somiglianza viene chiamato omologia. Generalizzando possiamo dire che gli arti anteriori dei vertebrati come del resto anche gli arti posteriori, mostrano lo stesso piano strutturale di base, sebbene siano adattati a funzioni molto diverse come il nuoto, il volo, la prensione degli oggetti; cioè diciamo che sono organi omologhi.
La spiegazione scientifica alla luce dell'evoluzione risulta molto semplice:i vertebrati discendono tutti da un antenato comune dal quale hanno ereditato queste strutture.
Omologie possono essere trovate a ogni livello dell'organizzazione della vita. Il pungiglione delle vespe è una modificazione dell'organo utilizzato da altri insetti per deporre le uova (è questo il motivo per cui pungono solo le femmine); le ali di una manta altro non sono che pinne pettorali enormemente sviluppate; gli sfiatatoi dei cetacei sono narici che si trovano nel posto più adatto per poter respirare mantenendo la testa sott'acqua.
Organi rudimentali
La presenza di organi rudimentali o vestigiali è un'ulteriore prova dell'evoluzione. Questi organi sono strutture che non compiono alcuna funzione ed appaiono perciò come assolutamente accessorie. Esse risultano però perfettamente omologhe con le stesse strutture ben sviluppate e funzionanti presenti in gruppi affini.
Boa e pitone, sebbene non abbiano gli arti posteriori, hanno, seppur fortemente ridotti, un femore e le ossa del bacino. Gli animali che vivono nelle caverne hanno generalmente occhi ridotti e assenti, forme vestigiali un tempo funzionali negli antenati che vivevano all'aperto.
Anche l'Uomo conserva alcuni organi vestigiali. L'appendice dell'intestino che probabilmente funzionava nei nostri lontani antenati come parte dell'apparato digerente, oggi non ha funzioni particolari e anzi spesso si infetta provocando l'appendicite e rendendo necessaria la sua asportazione. In corrispondenza dell'angolo sinistro dell'occhio umano permane una ridotta plica semilunare, la quale non è nient'alto che un rudimento della membrana nittitante che in altri Mammiferi come il cavallo, negli Uccelli, nei Rettili e negli Anfibi è molto più sviluppata e può essere abbassata come una tendina trasparente che protegge e lubrifica gli occhi senza impedire la visione.
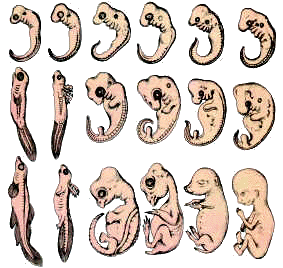
Prove embriologiche
Anche lo studio dello sviluppo embrionale fornisce
delle interessanti indicazioni e permette di intuire l'origine di organismi
diversi da antenati comuni.
Se si esaminano gli stadi dello sviluppo embrionale dei vertebrati, ci si
accorge che essi procedono seguendo un modello unico, a tal punto che se si
confrontano tra loro embrioni non molto avanzati di un pesce, di un anfibio
(salamandra), di un rettile (lucertola) e di alcuni mammiferi compreso l'Uomo,
ci si accorge che sono sorprendentemente simili tra loro e un embrione precoce
di anfibio è difficilmente distinguibile da un embrione precoce di mammifero.
In particolare in tutti gli embrioni citati compaiono le fessure branchiali, le
quali vengono mantenute allo stato adulto soltanto dai pesci nei quali hanno la
funzione di organi respiratori.
Questo fatto dimostra che tutti i vertebrati presi in considerazione hanno un
progenitore comune che doveva presentare respirazione di tipo branchiale.
Somiglianze nelle proteine
Alcune proteine sono presenti in tutti gli organismi viventi
e molte di esse presentano leggere differenze nella struttura nei diversi
gruppi di organismi. Le somiglianze e le differenze tra queste proteine
riflettono i rapporti di parentela tra le specie. Le proteine di un moscerino
sono più simili a quelle di una farfalla che a quelle di un cane. Inoltre c'è
maggiore somiglianza tra le proteine di un cane e quelle di una farfalla che
tra le proteine di questi animali e quelle di un tulipano. Queste osservazioni
confermano l'ide che un moscerino, una farfalla, un cane e un tulipano siano
tutti imparentati tra loro seppure con gradi diversi di parentela.
Prove biogeografiche
La biogeografia studia la distribuzione delle forme viventi
nelle varie regioni del globo e fornisce importanti prove a sostegno della
teoria dell'evoluzione. Se la distribuzione geografica delle specie non fosse
in qualche modo determinata dall'evoluzione dovremmo aspettarci di trovare una
determinata specie ovunque essa possa sopravvivere. Di fatto però, aree diverse
della Terra che pure hanno condizioni climatiche simili, come per esempio
l'Africa centrale e il Brasile sono abitate da animali e piante diverse. Le
specie che si sono evolute in Africa non hanno potuto estendere la loro
presenza in Brasile a causa della barriera naturale costituita dall'Oceano
Atlantico.
Regioni della Terra come l'Australia e la Nuova Zelanda, che sono state
separate dal resto del mondo in epoca molto antica, presentano flora e fauna
particolari.
In generale la distribuzione naturale degli organismi sulla Terra si può
comprendere solo facendo riferimento ai meccanismi evolutivi.

Selezione artificiale (allevamenti
e coltivazioni)
Nel corso di migliaia di anni l'uomo ha operato una
selezione artificiale che ha dato origine a nuove varietà di piante coltivate e
di animali di allevamento dotati di particolari caratteristiche utili. I
risultati di questa selezione artificiale forniscono una prova evidente che le
specie possono andare incontro a cambiamenti evolutivi in tempi relativamente
brevi.

Resistenza agli insetticidi e ai
farmaci
Un altro esempio di selezione naturale è lo sviluppo della
resistenza agli insetticidi. Sostanze chimiche velenose per gli insetti, come
il DDT, per un certo tempo hanno avuto molto successo, in seguito hanno perso
il loro prestigio non solo per la loro tendenza ad accumularsi nell'ambiente,
ma anche a causa dell'aumento straordinario di ceppi resistenti. Oggi almeno
255 specie di insetti sono resistenti a uno o più insetticidi.
Un esempio più recente di selezione naturale riguarda lo sviluppo della
resistenza dei batteri agli antibiotici.
I batteri resistenti sono delle varianti (mutanti) prodotte da un cambiamento
verificatosi nella popolazione iniziale e più adatte all'ambiente. I geni
batterici per la resistenza ai farmaci sono portati dai plasmidi,
piccole strutture contenenti DNA che possono essere trasferite da un batterio
all'altro.
La farfalla Biston betularia
Uno degli esempi più studiati di evoluzione per
selezione naturale è quello di Biston betularia, la farfalla delle
betulle molto comune in Inghilterra. La specie mostra due varianti di colore:
una forma chiara, dovuta ad un allele recessivo, e una forma scura (melanica),
dovuta a un allele dominante. Fino alla fine del secolo scorso, quasi tutti gli
esemplari erano di colore chiaro, utile per mimetizzarsi sulla corteccia chiara
delle betulle e sfuggire così agli uccelli predatori quando la corteccia delle
betulle era chiara. In seguito, lo sviluppo industriale e la conseguente
immissione di fuliggine nell'ambiente ha provocato il progressivo
annerimento dei tronchi; e di pari passo con il progredire dell'annerimento
dei tronchi, è aumentato drasticamente il numero degli esemplari scuri di
Biston a scapito di quelli chiari. Sulle cortecce scure, le varianti più
chiare risultavano più evidenti agli occhi dei predatori e venivano perciò
catturate più facilmente mentre le varianti più scure venivano avvantaggiate a
ogni generazione, così, a poco a poco, le forme scure sono diventate le più
diffuse e frequenti. Studi genetici hanno dimostrato che, in poco più di
cinquant'anni, la selezione naturale innescata dal cambiamento ambientale ha
provocato una variazione della frequenza dell'allele dominante da circa il 20%
a quasi il 100%.
Questa forma di "melanismo industriale" cioè il prevalere nelle zone
altamente industrializzate di forme scure, è piuttosto diffusa oggi in tutti i
paesi industrializzati e rappresenta un caso classico di selezione naturale.
Recentemente sono stati introdotti in Inghilterra rigidi controlli sul contenuto delle particelle di fumo e, grazie ad essi, il forte inquinamento da fuliggine è diminuito. La percentuale di farfalle chiare rispetto a quelle scure ha già ripreso ad aumentare.
|
Privacy |
Articolo informazione
Commentare questo articolo:Non sei registratoDevi essere registrato per commentare ISCRIVITI |
Copiare il codice nella pagina web del tuo sito. |
Copyright InfTub.com 2025