|
|
| |
GRUPPI FUNZIONALI
La chimica "organica" è la chimica dei composti del carbonio. Il carbonio (C) può formare 4 legami covalenti e nelle biomolecole lo troviamo insieme a ossigeno (O), azoto (N), idrogeno (H), fosforo (P) e zolfo (S).
Alcuni gruppi di atomi contenenti carbonio mostrano particolari e costanti proprietà chimiche, tali da poter essere identificati come "gruppi funzionali". Per analogia, alcuni gruppi di lettere (per esempio "c, h, e" oppure "g, l, i") formano, quando insieme, un suono particolare, che non è semplicemente la somma delle singole lettere).
I gruppi funzionali impartiscono caratteristiche chimico-fisiche particolari alle molecole che li contengono. Hanno tutti caratteristiche di polarità e possono formare ponti idrogeno sia con molecole d'acqua che tra loro.
Se c'è un solo gruppo funzionale nella molecola (il resto sono CH -CH ---), questo darà il nome alla molecola. Per molecole a 1, 2, 3 e 4 carboni, il nome deriva dal prefisso met-, et-, prop-, e but- seguito dalla desinenza che indica il tipo di funzione.
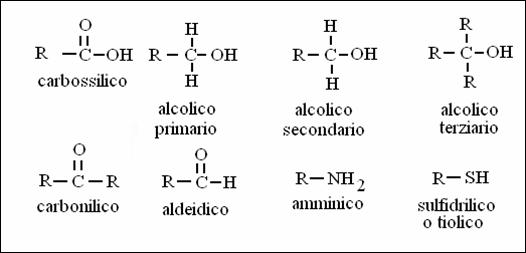
Acidi carbossilici: desinenza "oico" (o "ico nella vecchia nomenclatura), "ato" per la forma dissociata (es. acetato).
Esempi: HCOOH acido metanoico (formico)
CH -COOH acido etanoico (acetico: è l'acido dell'aceto)
CH -CH -COOH acido propanoico (propionico)
CH -CH -CH -COOH acido butanoico (butirrico)
Alcoli: desinenza "olo"
Esempi: CH OH metanolo
CH -CH OH etanolo (è il comune alcol per disinfezione)
CH -CH -CH OH propanolo
Chetoni: desinenza "one"
Esempio: CH -CO-CH propanone (acetone)
Aldeidi: desinenza "ale"
Esempi: HCOH metanale
CH -COH etanale
CH -CH -COH propanale
REAZIONI DEI GRUPPI FUNZIONALI
I gruppi funzionali sopra descritti hanno tendenza a reagire tra loro formando legami covalenti che possono unire tra loro molecole diverse o possono unire gruppi funzionali diversi sulla stessa molecola.
Queste sono le più importanti reazioni a cui partecipano e che rivestono grande importanza in biochimica.
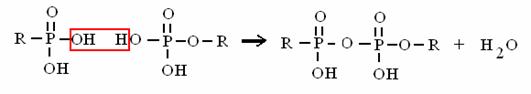
ACIDO + ALCOL ESTERE + ACQUA
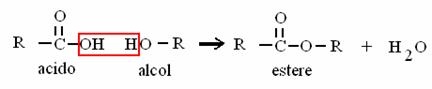
ACIDO FOSFORICO (dall'ATP) + ALCOL ESTERE FOSFORICO + ACQUA
è il legame del fosfato sul glucosio 6-P
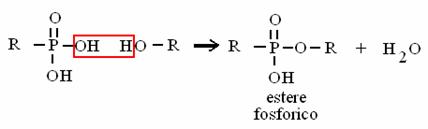
ACIDO + ACIDO ANIDRIDE + ACQUA
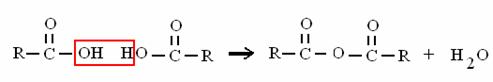
ACIDO FOSFORICO + ACIDO FOSF. ANIDRIDE FOSFORICA + ACQUA
è il legame ad alto contenuto energetico dell'ATP!
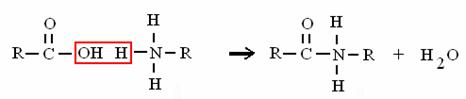
ACIDO + AMMINA AMMIDE + ACQUA
è il legame che unisce gli amminoacidi nelle proteine (legame peptidico)!
ALCOL + ALCOL ETERE + ACQUA
è il legame che unisce i monosaccaridi (legame glucosidico)!
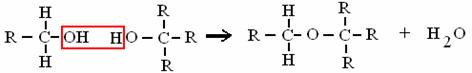
GR CARBONILICO (aldeide o chetone) + ALCOL EMIACETALE (NO ACQUA!)
è il legame intramolecolare nei monosaccaridi
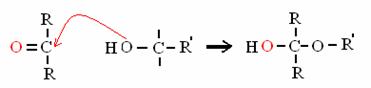
ACIDO + TIOLO (SULFIDRILE) TIOESTERE+ ACQUA
è il legame ad alta energia che attiva l'acil CoA e l'acetil CoA!
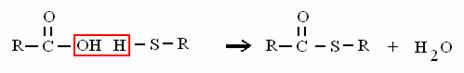
REAZIONI DI OSSIDO-RIDUZIONE
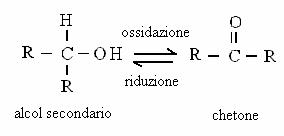
ALCOL PRIMARIO ALDEIDE ACIDO CARBOSSILICO
Alcoli, aldeidi e acidi carbossilici vengono trasformati gli uni negli altri attraverso reazioni di ossido-riduzione.
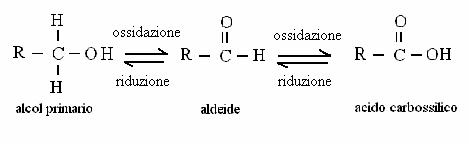
ALCOL SECONDARIO CHETONE
Tutte queste sono le reazioni del catabolismo ossidativo che producono energia sotto forma di coenzimi ridotti (NADH e FADH) secondo la reazione generale:
Xrid + coenzima Xox + coenzima ridotto
SULFIDRILE + SULFIDRILE DISOLFURO + equivalenti riducenti
è l'unico legame covalente nella struttura terziaria delle proteine
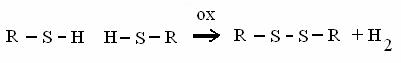
DG DI IDROLISI DI COMPOSTI FOSFORILATI
AD "ALTO CONTENUTO ENERGETICO"
DG (kcal/mole)
PEP -14.8
1.3 BPG -11.8
fosfocreatina - 10.3
ATP -7.3
Acetil CoA -7.5
Glucosio 6-P -3.3
Il potenziale di trasferimento del fosfato dell'ATP è intermedio tra quello di composti fosforilati donatori di fosfato (all'ADP) e quello di composti accettori di fosfato (dall'ATP).
BIOMOLECOLE
Sono tutti composti polifunzionali:
1. IDROSSIACIDI E CHETOACIDI
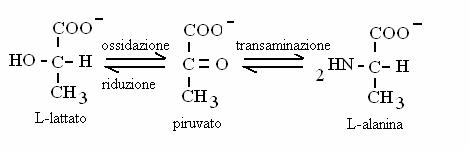
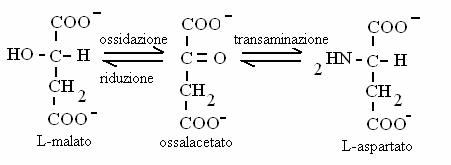
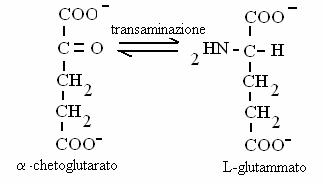
Contengono un gruppo carbossilico e un gruppo carbonilico (chetoacidi) oppure ossidrilico (idrossiacidi). Chetoacidi sono alcuni intermedi del ciclo di Krebs; idrossiacidi sono intermedi nella b-ossidazione degli acidi grassi. Chetoacidi e idrossiacidi possono trasformati gli uni negli altri attraverso reazioni di ossidoriduzione. Chetoacidi possono essere trasformati in amminoacidi (e viceversa) attraverso reazioni di transamminazione.
2. ZUCCHERI
comprendono: MONOSACCARIDI
DISACCARIDI (due monosaccaridi uniti insieme)
POLISACCARIDI (catena anche ramificata di monosaccaridi)
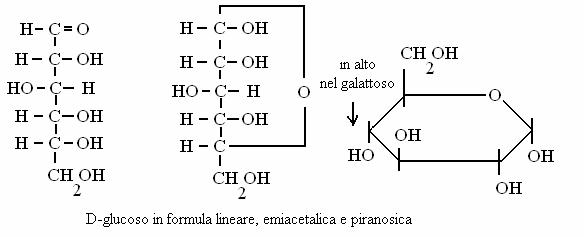
I MONOSACCARIDI contengono un gruppo carbonilico o un gruppo aldeidico insieme a uno o più gruppi ossidrilici. Si identificano quindi due famiglie di monosaccaridi: chetosi e aldosi.
ALDOSI (POLI-IDROSSIALDEIDI)
CHETOSI (POLI-IDROSSICHETONI)
I monosaccaridi più importanti in biochimica sono:
glucoso e galattoso (aldosi, esosi = 6 atomi di carbonio)
fruttoso (chetoso, esoso)
riboso (aldoso, pentoso = 5 atomi di carbonio)
Glucoso e fruttoso sono il principale carburante metabolico glucidico delle nostre cellule mentre il riboso partecipa alla costituzione degli acidi nucleici.
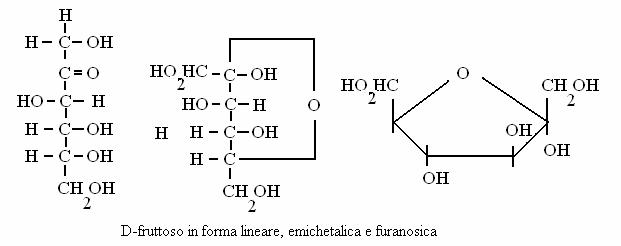
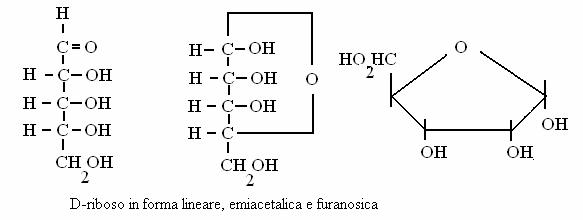
DISACCARIDI
Sono formati da due monosaccaridi, uniti con legame etere (tra 2 OH-) (legame glucosidico). I più importanti in biochimica umana sono:
SACCAROSO = GLUCOSO + FRUTTOSO (è lo zucchero di canna)
LATTOSO = GLUCOSO + GALATTOSO (è lo zucchero del latte)
MALTOSO = GLUCOSO + GLUCOSO (è lo zucchero del malto)
POLISACCARIDI
Sono formati dall'unione di molte migliaia di monosaccaridi. In natura hanno funzione di deposito di energia (glicogeno negli animali e amido nelle piante) oppure funzioni strutturali (di sostegno: cellulosa nelle piante, chitina negli insetti).
GLICOGENO = POLIMERO RAMIFICATO DEL GLUCOSO
Il glicogeno è il deposito di glucosio degli animali (solo in fegato e muscolo !).
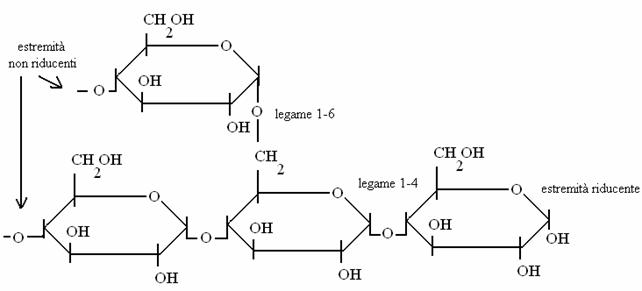
L'amido ha una struttura simile al glicogeno e gli enzimi animali che scindono il glicogeno sono attivi anche sull'amido (farina!). Invece la cellulosa ha legami 1-4 con una diversa configurazione spaziale dell'ossigeno e gli enzimi animali NON sono in grado di scinderla in glucosio (come fanno invece i ruminanti, grazie ai batteri intestinali!).
3. AMMINOACIDI
Contengono un gruppo amminico ed un gruppo carbossilico. Se il gruppo amminico è in posizione a rispetto al carbossile si chiamano a-amminoacidi e sono i costituenti delle proteine.
I 20 amminoacidi "fondamentali" (cioè quelli per cui c'è una tripletta codificante nel codice genetico) sono caratterizzati da un carbonio centrale (asimmetrico) cui sono legati: 1 gruppo amminico, 1 gruppo carbossilico, 1 idrogeno e la parte variabile nei 20 diversi amminoacidi fondamentali (indicata con R). Gli aa. nel nostro organismo sono solo isomeri L (come quello indicato in figura; l'isomero D avrebbe il gruppo amminico a destra). A seconda delle caratteristiche chimiche di R, gli aa. si classificano in 5 classi: polari, apolari, carichi, aromatici, solforati.
La grande varietà di gruppi R degli aa. conferisce alle proteine una grande versatilità chimica (capacità di interazione con gruppi funzionali diversi), e rende ragione del fatto che esse rappresentano le molecole "utensili" nelle cellule (con funzioni di catalizzatori, strutturali, di riconoscimento e difesa).
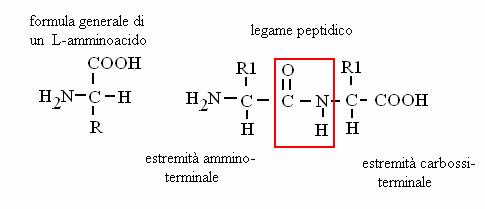
Le proteine sono polimeri di aa. uniti dal legame peptidico (un tipo di legame ammidico): tra gruppi peptidici di una stessa proteina che si vengono a trovare di fronte quando la proteina si ripiega si formano ponti idrogeno, che stabilizzano la struttura secondaria della proteina (a-elica o b-struttura).
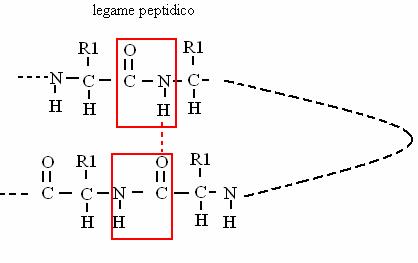
Le proteine sono polimeri lineari di aa. Si identificano 4 livelli strutturali, di complessità crescente.
La struttura primaria è la sequenza degli aa. uniti dai legami peptidici (covalenti), che si instaurano tra gruppo carbossilico e gruppo amminico di due aa. contigui.
La struttura secondaria può essere ad a-elica (elica destrogira con circa 4 aa per giro) o a b-struttura (anse che giacciono su un piano) ed è stabilizzata da ponti H tra gruppi peptidici lontani nella struttura primaria, ma che si vengono a trovare di fronte quando tratti discreti della proteina acquistano una struttura tridimensionale (cosa che succede già durante la sintesi proteica sui ribosomi).
La struttura terziaria è l'ulteriore ripiegamento dei tratti in a-elica e dei tratti in b-struttura della proteina a formare una struttura tridimensionale più complessa. Questa struttura è stabilizzata da legami deboli che si formano tra gruppi R di aa. che si vengono a trovare di fronte quando la proteina ha assunto la sua conformazione finale. Si tratta di legami ionici (tra gruppi R di aa. carichi positivamente e negativamente), ponti H ed interazioni idrofobiche (queste ultime sono in effetti il "motore" che guida il ripiegamento della proteina). L'unico legame covalente della struttura terziaria è il ponte disolfuro (tra due cisteine), presente non in tutte le proteine.
La struttura quaternaria è presente solo nelle proteine formate da più di una subunità: consiste nella posizione che assumono l'una rispetto all'altra le diverse subunità ed è stabilizzata dagli stessi tipi di legame descritti per la struttura terziaria (esclusi i ponti disolfuro) che si instaurano tra guppi R di aa. su catene proteiche diverse.
4. LIPIDI
I lipidi più importanti in biochimica umana sono:
ACIDI GRASSI
TRIGLICERIDI
FOSFOGLICERIDI
COLESTEROLO
ACIDI GRASSI = acidi carbossilici a lunga catena (C16-C18) saturi o insaturi (senza o con doppi legami tra i carboni)
![]()
Gli acidi grassi sono la fonte primaria (in termini quantitativi) di energia per il nostro organismo e sono presenti soprattutto sotto forma di trigliceridi (vedi sotto), cioè esterificati (legati con legame estere) al glicerolo (un polialcol).
Noi umani non siamo in grado di sintetizzare acidi grassi con doppi legami vicini al metile terminale (w 3): questi acidi grassi insaturi sono detti "essenziali" e vanno assunti con la dieta.
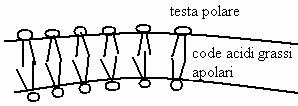
I saponi sono acidi grassi a lunga catena salificati con Na+. I saponi in acqua formano strutture sopramolecolari dette "micelle" (delle microscopiche gocce): le code apolari degli acidi grassi, che fuggono l'acqua, sono rivolte verso il centro della micella mentre la testa polare (il gruppo carbossilico dissociato -COO ) è rivolto verso l'acqua perché è carico (interagisce con gli H parzialmente positivi dell'acqua). Quindi il centro della micella è apolare. Sostanza lipofile (grassi) tenderanno a "sciogliersi" dentro la micella (il sapone "scioglie" la macchie d'unto!).
TRIGLICERIDI = esteri del glicerolo con 3 acidi grassi
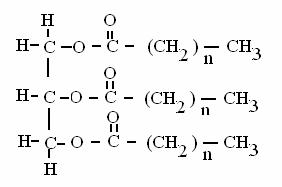
I trigliceridi sono i grassi di deposito del nostro organismo, accumulati nel tessuto adiposo, dentro gli adipociti e sono la principale forma di deposito di energia metabolica del nostro organismo (120000 kcal totali contro le 1200 del glicogeno). Il tessuto adiposo è localizzato nel sottocute e intorno agli organi interni. L'idrolisi dei trigliceridi ad acidi grassi e glicerolo è catalizzata dagli enzimi lipasi. I trigliceridi non si sciolgono in acqua (non hanno alcuna parte polare nella molecola), ma viaggiano nel sangue ricoperti da una guscio proteico, sotto forma di chilomicroni e di particelle lipoproteiche (VLDV, LDL, HDL).
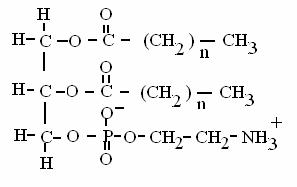
FOSFOGLICERIDI = esteri del glicerolo con 2 acidi grassi e una "testa polare" contenente fosfato (da cui il nome fosfogliceridi) che può essere fosfo-etanolammina (in figura), fosfo-serina, fosfo-inositolo o fosfo-colina. Insieme al colesterolo, sono i costituenti delle membrane cellulari. Formano strutture sopramolecolari chiamate doppi strati (non formano micelle).
COLESTEROLO = struttura a 4 anelli carbociclici condensati (a ogni angolo e su ogni punta c'è un carbonio). Il colesterolo è un componente importante delle membrane cellulari e dal colesterolo il fegato produce i sali biliari (importanti per la digestione).
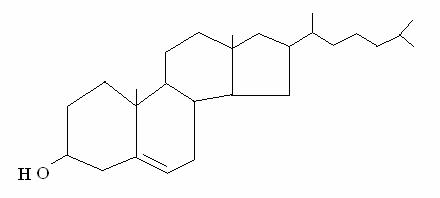
L'unica parte polare della molecola è l'OH: nei doppi strati delle membrane l'ossidrile si orienta verso la fase acquosa (intra- o extra-cellulare) e il resto della molecola (che è planare) si infila in mezzo alle code degli acidi grassi. Il colesterolo impedisce l'impaccamento delle code degli acidi grassi e assicura fluidità alle membrane.
5. BASI PURINICHE E PIRIMIDINICHE
Le basi puriniche e pirimidiniche sono costituenti dei nucleotidi. Nella cellula, i nucleotidi hanno funzioni importanti: energetiche (ATP è la moneta corrente per tutti gli scambi energetici della cellula), formano parte della molecola dei coenzimi NAD e FAD (i trasportatori di elettroni nelle reazione di ossido-riduzione del catabolismo ossidativo) e sono le unità costitutive degli acidi nucleici (DNA e RNA).
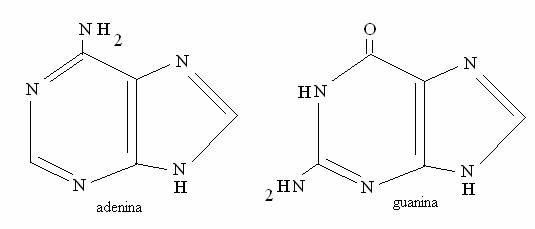
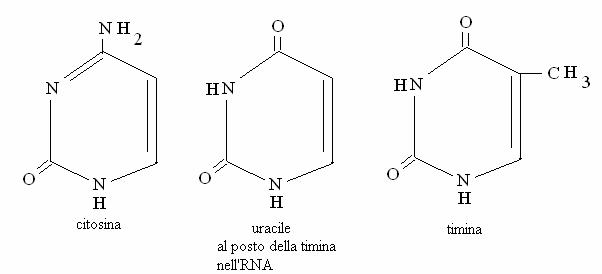
Le basi puriniche sono fromate da 2 anelli condensati, mentre le pirimidiniche hanno un solo anello. Sono dette "basi" perché alcuni azoti possono protonarsi (diventare H+) sottraendo H+ dalla soluzione.
Basi puriniche (A e G)
Basi pirimidiniche (C, U, T)
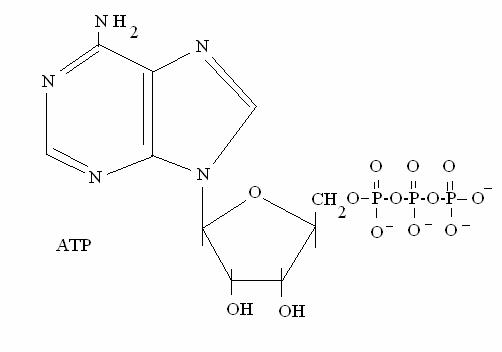
NUCLEOSIDE = BASE + RIBOSO
NUCLEOTIDE = BASE + RIBOSO + FOSFATO (mono-, bi- o trifosfato)
Adenosina trifosfato (ATP)
Un esempio di nucleotide trifosfato è l'ATP (adenosine triphosphate): l'ATP è la "moneta corrente" per tutti i processi che producono e che consumano energia nella cellula. E' caratterizzato da un alto potenziale di trasferimento del fosfato: cioè l'idrolisi del legame anidridico che unisce tra loro gli ultimi 2 fosfati, libera tanta energia:
ATP + H2O ADP + Pi (fosfato inorganico) DG = - 7.5 kcal/mole
I motivi sono chimici: la repulsione tra cariche negative sui fosfati (minore quando perde un fosfato) destabilizza la molecola dell'ATP (= alto contenuto energetico potenziale) e la stabilizzazione di risonanza del fosfato (una volta idrolizzato) spinge la reazione verso l'idrolisi.
L'idrolisi dell'ATP viene sfruttata a volte nel metabolismo per spostare l'equilibrio di una reazione energeticamente sfavorevole:
A B DG = + 4 kcal/mole
ATP + H2O ADP + Pi (fosfato inorganico) DG = - 7.5 kcal/mole
A + ATP + H2O B + ADP + Pi DG = - 3.5 kcal/mole
COENZIMI VITAMINICI
Gli enzimi sono i catalizzatori delle reazioni biologiche: molto spesso però gli enzimi da soli non sono in grado di catalizzare la loro specifica reazione, ma necessitano di molecole, dette coenzimi, che li "aiutino". Il coenzima interviene con alcuni suoi gruppi funzionali specifici, che l'enzima non possiede, e che sono necessari durante la catalisi.
I coenzimi vengono sintetizzati nelle nostre cellule a partire da molecole, le vitamine idrosolubili, che invece non siamo in grado di produrre: in altre parole, i coenzimi sono vitamine modificate chimicamente. Le vitamine sono prodotte dalle piante e, in qualche caso, dai batteri intestinali.
La necessità di una dieta contenete frutta e verdure fresche era già stata riconosciuta nel '700, osservando gli effetti nefasti dei lunghi viaggi in mare (dieta a base di cereali secchi e carne salata) sullo stato di salute dei marinai.
Non tutte le vitamine hanno una funzione coenzimatica: le vitamine liposolubili (A,D,E e K) sono invece delle molecole segnale, da considerarsi simili a ormoni per la molteplicità dei loro effetti.
I sintomi da deficit vitaminico sono rari nella nostra società, ma possono verificarsi in: soggetti anziani (cattiva alimentazione, malassorbimento), alcolisti (cattiva alimentazione, epatopatia, malassorbimento), terapia anticonvulsivante cronica (deficit folati e vit. D), ripetute gravidanze con allattamento (aumentato fabbisogno), gastrite atrofica (deficit produzione fattore intrinseco necessario per assorbimento B12), diete vegetariane strette e di lunga durata.
I sintomi generici da deficit sono simili per tutte le vitamine coenzimatiche, poiché derivano dall'alterazione del metabolismo energetico, che colpisce prevalentemente i tessuti con elevate necessità (in rapida crescita, come epiteli, mucose, cellule del sangue, o ad attività metabolica elevata, come sistema nervoso e apparato muscolare). I sintomi generici comprendono dermatite, cheilite, glossite, neuropatie periferiche, incoordinazione motoria, faticabilità, malessere, diarrea.
|
VITAMINA |
COENZIMA |
ENZIMA/FUNZIONE |
SINTOMI DA DEFICIT |
RDA mg |
|
Tiamina (B1) |
TPP |
Piruvato deidrogenasi e KGA-deidrogenasi (ciclo di Krebs) |
Atassia, oftalmoplegia, confusione mentale, faticabilità. |
|
|
Ac. Pantotenico (B5) |
Coenzima A |
Trasportatore-attivatore di acili |
Non noti, forse perché molto diffuso negli alimenti. |
|
|
Piridossina (B6) |
PLP |
Transaminasi |
Neuropatie periferiche, anemia sideroblastica. |
|
|
Biotina |
Biocitina |
Carbossilasi |
Prodotta dai batteri intestinali. |
|
|
Ac. folico |
THF |
Trasf. unità carboniose nella sintesi purine e dTMP (sintesi DNA) |
Anemia megaloblastica; difetti del tubo neurale in neonati da madri deficitarie. |
|
|
Ac. nicotinico |
NAD+ |
Deidrogenasi (trasf. elettroni) |
Pellagra: dermatite fotoreattiva, diarrea, demenza |
|
|
Riboflavina (B2) |
FAD |
Deidrogenasi (trasf. elettroni) |
Cheilite, dermatite, glossite |
|
|
Ac. Ascorbico (C) |
Non è un coenzima, ma è necessaria come substrato durante la sintesi del collagene. Aiuta l'assorbimento del ferro. |
Idrossilazione di Lys e Pro nel proto-collagene, necessario per legami covalenti tra fibrille. Anti-ossidante. |
Scorbuto: fragilità capillare e del collagene tissutale, osteoporosi, cattiva riparazione ferite |
|
|
Cobalamina (B12) |
Non è un coenzima, ma è necessaria durante una reazione del THF. Nella carne (fegato). |
Recupero del THF dal metil-THF; recupero del metilmalonil-CoA a succinil-CoA. |
Anemia megaloblastica; deficit neurologici da demielinizza-zione. |
|
RDA = recommended dietary allowance = dose giornaliera consigliata
VITAMINE LIPOSOLUBILI
Le vitamine liposolubili non formano coenzimi, ma sono piuttosto da considerare come molecole segnale (ormoni). Sono depositate nel fegato: i sintomi da deficit possono richiedere anni prima di manifestarsi a seguito di condizioni favorenti: malassorbimento lipidico, epatopatie croniche, alcolismo, terapia antibiotica (K), nefropatie croniche (K).
|
VITAMINA |
FUNZIONE |
SINTOMI DA DEFICIT |
RDA mg |
|
A retinolo, retinale, ac. retinico |
Anti-ossidante, regola proliferazione e differenziamento cellulare, pigmento visivo (rodopsina) |
Cecità notturna, ipercheratosi, anemia. |
b-carotene) |
|
D colecalciferolo, D3 7-deidrocol. è attivato a D3 dalla luce UV. D3 è attivato per idros-silazione nel fegato e nel rene a 1,25-(OH)2D. |
Stimola assorbimento intestinale del Ca2+, inibisce l'escrezione renale e aumenta il riassorbimento di Ca2+ dall'osso (insieme a PTH) |
Rachitismo nei bambini, osteomalacia nell'adulto |
|
|
E Miscela di composti vegetali: tocoferoli |
Anti-ossidante nelle membrane, lipoproteine e tessuto adiposo; stabilizza CoQ (respirazione mitocon.) |
Non noti |
|
|
K Sintetizzato dai batteri intestinali |
Richiesta per la sintesi epatica dei fattori della coagulazione 2°, 7°, 9° e 10°, e di osteocalcina |
Aumento del tempo di coagulazione; osteoporosi. |
|
|
Privacy |
Articolo informazione
Commentare questo articolo:Non sei registratoDevi essere registrato per commentare ISCRIVITI |
Copiare il codice nella pagina web del tuo sito. |
Copyright InfTub.com 2025