|
|
| |
La divisione dei muscoli
A livello tissutale gli apparati contrattili possono risultare composti da due tipi di tessuto ben distinti l'uno dall'altro: il muscolo liscio e quello striato.· Il tessuto muscolare liscio. É detto così perché, osservato al microscopio, non presenta alcuna evidenza di striature trasversali (caratteristiche invece del muscolo striato). Il muscolo liscio si trova a livello viscerale ed è responsabile dei movimenti delle strutture non accessibili al controllo diretto della coscienza (ad esempio della peristalsi intestinale, della contrazione o del rilassamento delle pareti delle arterie, etc.). Il muscolo liscio si t 646f58g rova praticamente nella parete di ogni struttura cava dell'organismo; il suo stato di contrazione o di rilassamento determina lo stato di rigidità della parete stessa.· Il tessuto muscolare striato. É caratteristico degli apparati motori sottoposti al diretto controllo da parte della volontà, tipicamente della muscolatura scheletrica. Il termine ""striato"" deriva dall'aspetto che il tessuto, preparato su di un vetrino in sezione sottile, assume se osservato al microscopio: si osserva la presenza di un regolare susseguirsi di bande alternativamente chiare o scure che determinano la presenza di una vera e propria ""striatura"". La bandatura del muscolo striato è dovuta al particolare ordine con il quale sono disposti componenti molecolari della struttura contrattile. Nel muscolo liscio questi componenti sono disposti con un certo disordine strutturale.
I muscoli scheletrici
Le fibre muscolari scheletriche sono elementi citologici molto allungati e del tutto particolari per la struttura e le caratteristiche funzionali e sono riunite in fasci di sempre maggiori dimensione da sepimenti connettivali che costituiscono un "impalcatura" (endomisio) cui le fibre sono ancorate. L'endomisio, agli estremi del muscolo, si unisce col perimisio (la lamina connettivale che avvolge tutto il muscolo) ed insieme si continuano nei tendini. Nel connettivo interposto tra le fibre decorrono pure i nervi, costituiti da fibre motorie e sensitive.
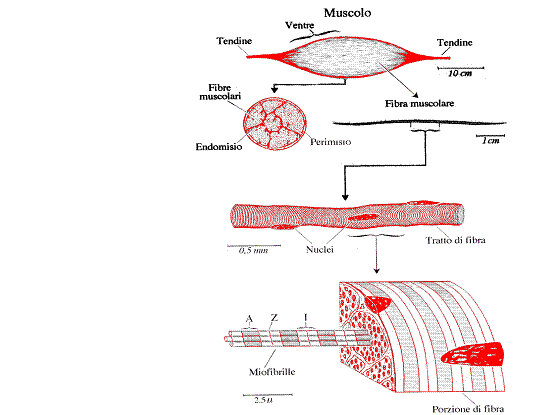
Le fibre muscolari scheletriche sono elementi cellulari non solo molto allungati, ma di dimensioni eccezzionalmente grandi. Ogni fibra muscolare è delimitata da una membrana: il sarcolemma .Al suo interno si trova il sarcoplasma contenenti numerosi nuclei. Il volume di ogni fibra è quasi interamente occupato dall'apparato contrattile, costituito dalle miofibrille.
All'interno delle fibre è inoltre presente una rete di tubuli intracellulari, il reticolo sarcoplasmatico , che in parte comunica con l'esterno e che corrisponde al reticolo endoplasmatico.
Il sarcoplasma è compreso quasi tutto in un sottile strato disposto sotto il sarcolemma. E' ricco di proteine, di glicogeno e dei fosfati altamente energetici: l'adenosintrifosfato (ATP) ed il creatinfosfato (CP). Si trovano anche, particolarmente numerosi, i mitocondri, che si distribuiscono anche negli spazi tra le miofibrille.
I nuclei. La loro abbondante presenza testimonia che le fibre dei muscoli scheletrici originano embriologicamente da elementi progenitori (i mioblasti) nei quali ripetute moltiplicazioni nucleari non sono seguite da altrettante divisioni del corpo cellulare; una fibra muscolare scheletrica ha perciò il carattere di un plasmodio.
Le miofibrille sono l'apparato contrattile delle fibre muscolari. Lungo
ogni miofibrilla si replica, per l'intera lunghezza, un alternanza di zone mono
e bi-rifrangenti. La sequenza delle zone con diversa rifrangenza è dovuta al
ripetersi di unità ultrastrutturali sempre uguali: i sarcomeri. Ogni sarcomero
è delimitato da due netti sepimenti disposti a metà della zona monorifrangente:
le strie Z . La zona monorifrangente è denominata zona I (isotropa), quella
birifrangente è denominata zona A (anisotropa). Al centro della zona A vi è una
zona otticamente meno densa, la zona H, che scompare quando la fibra è
contratta. 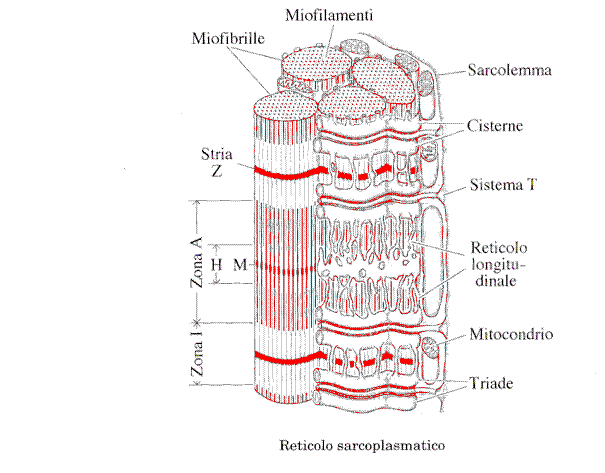
Le caratteristiche dei sarcomeri sono dovute alla presenza in essi di una serie ordinata di miofilamenti orientati parallelamente all'asse delle miofibrille. Ci sono i miofilamenti sottili (actina,tropomiosina, troponina) e quelli spessi (miosina). Ove sono presenti entrambi essi sono disposti secondo una simmetria esagonale; ogni miofilamento spesso è infatti circondato regolarmente da 6 miofilamenti sottili ed ogni miofilamento sottile è circondato da 3 miofilamenti spessi.
Per demolizione enzimatica la molecola miosinica si risolve in 3 coppie di catene polipeptidiche uguali a due a due ma di diversa dimensione: una prima coppia costituisce la coda, ed in essa le due catene polipeptidiche sono avvolte a doppia elica; la porzione terminale di ogni catena pesante si avvolge in struttura terziaria e va a formare gran parte di una delle due teste; la altre due coppie vanno invece a formare il tratto di connessione delle "teste" con la"coda" della molecola. Il tratto di congiunzione con le teste può flettersi sulla parte lineare e costituisce una sorta di "braccio". Le molecole miosiniche si orientano in modo contrapposto e secondo un andamento a spirale . Lungo il miofilamento sporgono così su uno stesso piano ortogonale al suo asse, 3 duplici teste" miosiniche ruotate di 120 l'una rispetto all'altra, che formano una "corona"; ogni corona è ruotata di 60 rispetto alla precedente. Ne viene che da ogni metà del miofilamento, sporgano, nel senso della lunghezza, 6 file di "teste" distanziate di 430 Angstrom. Questa disposizione fa si che ogni fila di teste venga affrontata, con simmetria esagonale, ad uno dei 6 miofilamenti sottili che circondano ogni miofilamento spesso.
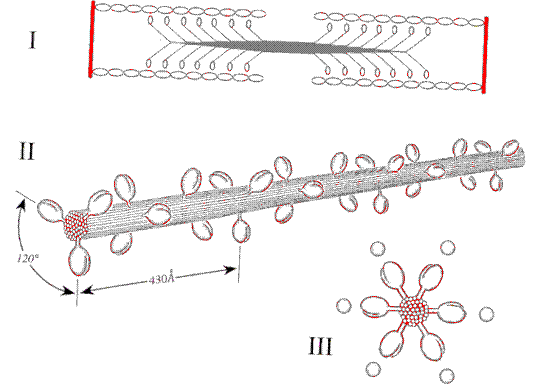
I mioflamenti sottili sono costituiti da due lunghe catene di actina filamentosa avvolte a spirale. Ogni molecola globulare di actina presenta un sito di legame capace di combinarsi con la testa della miosina. La tropomiosina costituisce un subfilamento continuo. Ciascuna molecola troponinica invece è composta da tre subunità peptidiche. La prima (T) è sede del legame con la tropomiosina, la seconda (I) è implicata nel controllo dei siti di legame dell'actina, la terza (C) ha affinità per gli ioni Ca2+.
Nel reticolo sarcoplasmatico si distinguono due parti: un sistema tubulare trasverso che comunica con l'esterno mediante una serie di pori e un sistema longitudinale ai cui estremi presenta dilatazioni dette "cisterne". Nelle fibre muscolari scheletriche dei Mammiferi vi sono due sistemi trasversi per ogni sarcomero.
-Eccitabilità delle fibre muscolari scheletriche-
In condizioni fisiologiche le fibre vengono portate in contrazione dalle fibre nervose motorie tramite le giunzioni sinaptiche neuromuscolari o placche motrici. Il potenziale d'azione viene innescato dal potenziale postsinaptico delle placche motrici (potenziale di placca). Mentre la successione delle fasi è simile a quella del potenziale d'azione delle fibre nervose, la durata totale del potenziale d'azione delle fibre muscolari è notevolmente maggiore (2-5 msec). Anche nelle fibre muscolari il potenziale d'azione si propaga da luogo d'insorgenza lungo l'intera fibra tramite le correnti elettrotoniche. Anche la contrazione viene a manifestarsi come un onda che segue, con un breve ritardo, il potenziale d'azione ma ha, rispetto a questo, una durata molto maggiore.
La fibra riacquista la sua normale eccitabilità prima che inizi la contrazione, ed è perciò in grado di rispondere, se nuovamente stimolata, anche se sta già contraendosi. Questo per esempio non avviene nelle fibrocellule miocardiche.
-Il meccanismo della contrazione dei sarcomeri-
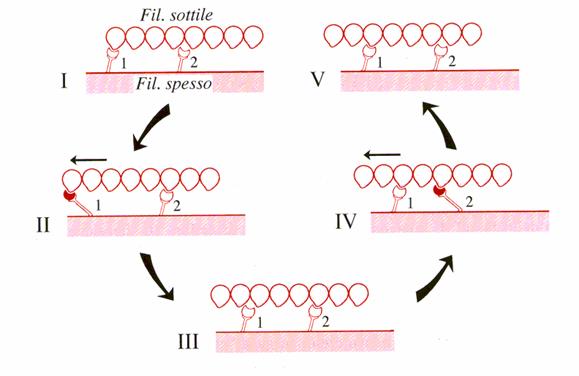
Il ciclo deve ripetersi con velocità molto elevata in ogni "ponte".
Per spiegare la continuità dello scorrimento si ammette che gruppi di ponti entrino in azione in modo desincronizzato. La desincronizzazione sarebbe assicurata dal fatto che ogni giro della doppia spirale actinica non coincide con la distanza delle "teste" di miosina della "fila" rivolta verso di esso.
-Accoppiamento elettro-meccanico-
Al seguito di un singolo potenziale d'azione si susseguono, nella fibra muscolare, una fase di contrazione e, con breve ritardo, una fase di rilasciamento che riporta la fibra nelle condizioni di riposo. L'accoppiamento è mediato da ioni Ca2+. Questi ioni provengono dal reticolo sarcoplasmatico longitudinale dove sono sequestrati in forma legata. In particolare vengono accumulati nelle cisterne. Il passaggio del potenziale d'azione lungo la fibra muscolare è immediatamente accompagnato dal liberarsi di calcio dalle cisterne del reticolo longitudinale.
Lo stabilirsi di un legame tra le teste miosiniche e le molecole actiniche, in sé, non è Calcio-dipendente. E' necessario ammettere la presenza di un processo che renda Calcio-dipendente la loro interazione. I subfilamenti di tropomiosina sono disposti nei solchi della doppia spirale actinica dei filamenti sottili in modo da "mascherare" i "siti di legame". Il legame del Calcio con la subunità C della troponina ne modifica la conformazione e fa spostare la tropomiosina smascherando i siti di legame. Il sistema troponina-tropomiosina è insomma un sistema inibitore dell'apparato contrattile. Gli ioni Calcio dopo che hanno esplicato la loro funzione vengono allontanati: o espulsi dal sarcolemma o riassorbiti nel reticolo. In entrambi casi il meccanismo è operato dalla pompa ionica del Calcio. L'efficienza della pompa non è uguale nei diversi tipi di fibra muscolare e da essa dipende la durata della contrazione che segue un potenziale. Lo stato attivo dell'apparato contrattile, vale a dire l'intensità e la durata della contrazione dipende dalla quantità di ioni rilasciati ad ogni impulso e dal tempo per cui gli ioni restano liberi di agire.
Le proteine miofibrillari hanno proprietà ATPasiche tanto che il complesso actomiosina in vitro in presenza di ATP dà un superprecipitato. Esperimenti più approfonditi hanno dimostrato che solo la miosina possiede la proprietà di un enzima ATPasico e la esplica a livello delle teste.Quando la miosina è in presenza di actina questa attività viene esaltata e diventa continua.
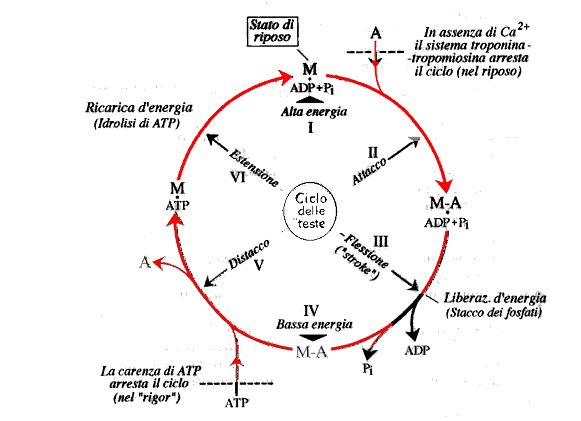
-Il ciclo operativo dei "ponti"e l'idrolisi dell'ATP-
E' indubbio che la fase efficace, quella cioè in cui l'energia di origine chimica si converte in energia meccanica è una modificazione strutturale delle "teste" di miosina. Tutavia vi sono prove sicureche la reazione defosforilativa ATP ADP + Pi non si verifica nella fase di "stroke", ma successivamente quando le teste miosiniche si staccano dalle molecole di actina e ritornano nella configurazione atta a iniziare un nuovo ciclo.
Come si può spiegare questo?
1- sulle teste miosiniche dobbiamo avere un sito ATP-asico ed un sito actinico.
2- il sito ATP-asico può essere indifferentemente occupato dall'intero ATP o dall' ADP+Pi.
- sul sito actinico può esserci o no legata l'actina.
3- i due siti si influenzano reciprocamente
|
Il complesso M-ADP+Pi ha grande affinità |
|
Il complesso M-ATP non ha affinità per l'actina |
|
per converso --> la combinazione con l'actina riduce l'affinità per ADP+Pi e aumenta quella per l'ATP. |
Ciclo idrolitico dell'ATP
I muscoli del torace
I muscoli pettorali, chiamati anche muscoli dei gran pettorali, fanno parte della categoria dei cosiddetti muscoli grandi e si dividono in tre fasce: pettorali alti, fascia centrale e pettorali bassi che si dividono a loro volta in fasce interne ed in fasce esterne.
Trattandosi di muscoli grandi e molto complessi, vanno allenati con una certa cura ed intensità, privilegiando esercizi che lavorino meglio le fasce più carenti, pur essendo vero che la maggior parte degli esercizi per i pettorali lavorano comunque tutto il muscolo, pur battendo di più su una fascia piuttosto che su un'altra.
Generalmente i pettorali si allenano il primo giorno utile di allenamento della settimana (che in genere coincide con l'inizio della settimana stessa), accoppiandoli con i bicipiti o con i tricipiti in fase di massa, piuttosto che con le spalle (deltoidi) per poche sedute di allenamento dopo Natale, o ancora con i dorsali in fase di definizione, dopo Pasqua fino all'estate.
I muscoli del addome
I muscoli addominali, detti anche muscoli del retto dell'addome, fanno parte della categoria dei muscoli grandi del corpo, ma nonostante ciò, sono muscoli molto delicati e vanno allenati con cura. Si dividono in tre fasce muscolari: addominali alti, addominali bassi e addominali laterali, detti anche addominali obliqui.
Gli addominali alti sono costituiti da 6 "quadrati", disposti in fila per due su tre livelli; gli addominali bassi sono composti da una sola fascia che inizia dall'altezza dell'ombelico e si congiunge col basso ventre, mentre gli addominali laterali ricoprono le costole e ne assumono la stessa forma, una volta ben sviluppati e privi di strati di grasso.
Si consiglia l'allenamento costante almeno due o tre volte a settimana, seguendo un programma di allenamento variabile a seconda della propria corporatura, delle proprie esigenze e della scheda di allenamento che si sta seguendo.
Si consideri che la difficoltà nell'avere degli addominali scolpiti non dipende solo dal tipo di allenamento svolto, ma anche da quanto strato di adipe (grasso) vi è in accumulo in questa particolare zona del corpo, molto soggetto all'accumulo: è quindi necessario seguire una buona dieta e svolgere attività aerobica al fine di poter godere della loro visibilità.
I muscoli dell'arto superiore
I muscoli dell'arto superiore si distinguono in
intrinseci ed estrinseci.
I muscoli estrinseci prendono inserzione sullo scheletro dell'arto superiore,
ma hanno la loro origine al di fuori di questo; sono i muscoli
spinoappendicolari e toracoappendicolari.
I muscoli intrinseci presentano sia l'origine sia l'inserzione sullo scheletro
dell'arto. In direzione prossimo-distale, essi si distinguono in:
I muscoli dell'arto inferiore
muscoli dell'arto inferiore si suddividono in quattro gruppi che, in direzione prossimo-distale, sono: i muscoli dell'anca, i muscoli della coscia, i muscoli della gamba e i muscoli del piede.
I muscoli dell'anca si distinguono in:
Le fasce dei muscoli dell'anca sono rappresentate dalla fascia iliaca e dalla fascia glutea (o fascia dei muscoli grande e medio gluteo).
I muscoli della coscia si distinguono in:
I muscoli della gamba si distinguono in:
Essi si trovano in una loggia delimitata dalla membrana interossea della gamba e dai margini anteriori della tibia e della fibula.
I muscoli del piede si distinguono in:
Il tono muscolare
Per tono muscolare intendiamo il costante stato di contrazione dei muscoli in una data posizione (seduti, in piedi, ecc.); il suo aumento dipende dalla trasmissione nervosa che avviene attraverso cervello e midollo spinale.
MECCANISMO GENERALE DELLA CONTRAZIONE MUSCOLARE
L'avvio e il corso della contrazione muscolare hanno luogo secondo questa sequenza di eventi:
1. Un potenziale d'azione viaggia lungo una fibra nervosa motrice fino alle sue terminazioni su un certo numero di fibre muscolari.
2. A livello di ciascuna terminazione, la fibra nervosa secerne una piccola quantità di una sostanza neurotrasmettitrice, l'acetilcolina.
3. L'acetilcolina agisce su un'area circoscritta della membrana della fibra muscolare determinando l'apertura di molteplici canali ionici a controllo di ligando (colinergici nicotinici) presenti nella membrana stessa.
4. L'apertura di questi canali permette l'ingresso di grandi quantità di ioni sodio all'interno della membrana della fibra muscolare nel punto in cui termina la, fibra nervosa.
5. Si genera così a questo livello un potenziale d'azione che si propaga lungo la membrana della fibra muscolare allo stesso modo in cui un potenziale d'azione viaggia lungo la membrana di fibre nervose.
6. Il potenziale d'azione depolarizza la membrana della fibra muscolare e si propaga anche in profondità nella fibra stessa. Qui provoca la liberazione dal reticolo sarcoplasmatico nelle miofibrille di grandi quantità di ioni calcio immagazzinati nello stesso reticolo.
7. Gli ioni calcio innescano un processo che dà origine a forze di attrazione tra i filamenti di actina e quelli di miosina che slittando gli uni sugli altri determinano la contrazione della fibra muscolare.
8. Dopo una frazione di secondo, gli ioni calcio vengono richiamati all'interno dei reticolo sarcoplasmatico, dove rimangono immagazzinati sino a che la fibra muscolare non è investita da un nuovo potenziale d'azione, e la contrazione muscolare cessa.
Proprietà meccaniche
Classici preparati sperimentali usati per indagare le proprietà meccaniche dei muscoli sono i muscoli gastrocnemio o sartorio di rana. Il sartorio è connesso ad una estremità al pelvi e all'altra estremità alla tibia a livello del ginocchio. La sua contrazione muove la gamba in avanti e la flette all'altezza del ginocchio. Il muscolo può essere agevolmente isolato e i tendini possono essere facilmente fissati ad opportuni apparati di registrazione.
Abbiamo visto in precedenza che le fibre muscolari sono eccitabili; un muscolo, quindi, può essere stimolato mediante applicazione diretta di stimoli elettrici o attraverso la stimolazione del nervo motore.
Quando un muscolo si contrae esso esercita una forza su ciò a cui è attaccato (questa forza è uguale alla tensione nel muscolo) e si accorcia se gli è consentito di farlo. Pertanto noi possiamo misurare due diverse variabili durante la contrazione di un muscolo: la sua lunghezza e la sua tensione. Nella maggior parte dei casi una di queste due variabili è mantenuta costante durante la contrazione. Nella contrazione isometrica al muscolo non è consentito l'accorciamento (la sua lunghezza è mantenuta costante) e si misura la tensione prodotta. Nella contrazione isotonica il carico sul muscolo (che è uguale alla tensione prodotta nel muscolo) è mantenuto costante e si misura la accorciamento.
L'acido lattico
Dobbiamo fare una premessa, come avviene la contrazione muscolare? L'ATP
(acido adenosinotrifosforico), presente in piccole quantità nei muscoli, può
cedere costantemente energia grazie al suo continuo riformarsi in quanto
cedendo energia ai muscoli perde un radicale fosforico diventando ADP (acido
adenosindifosforico) che a sua volta riacqista velocemente energia ridiventando
ATP. Perchè si verifichi questo continuo processo abbiamo bisogno di sfruttare
una fonte di energia che può essere ottenuta in due modi:
1.con.l'aiuto.dell'ossigeno.(processo.aerobico)
2.senza.l'aiuto.dell'ossigeno.(processo.anaerobico)
Nel primo caso, per creare energia, vengono bruciati gli zuccheri (glicogeno)
presenti nei muscoli e all'occorrenza gli acidi grassi, questo processo di
combustione ha bisogno di ossigeno e produce l'energia necessaria per
trasformare l'ADP in ATP, lasciando come prodotto di rifiuto anidride carbonica
e acqua che vengono velocemente eliminate tramite il sangue di ritorno dai
muscoli, quindi con l'espirazione ed il sudore; quindi
zucchero
-------------+ossigeno=energia+CO2+H2O
ograssi
ADP+energia=ATP
quando però i muscoli sono sottoposti a sforzi intensi e rapidi, essi sfruttano
una notevole quantità di ATP destinato a diventare ADP, di conseguenza
l'ossigeno che viene inspirato non è più sufficiente a ritrasformare
velocemente l'ADP in ATP, ed a questo punto entra in funzione il 2° meccanismo
per produrre energia.
Il processo anaerobico può, a sua volta, essere di 2 tipi:
- processo anaerobico alattacido dove lo sforzo muscolare non produce acido
lattico in quanto lo sforzo stesso è di brevissima durata;
- processo anaerobico lattacido dove gli zuccheri scindendosi producono energia
e acido lattico. Una parte si accumula nei muscoli, un'altra viene lentamente
portata dal sangue al fegato che provvede a trasformarlo in zucchero, quindi:
zucchero=energia+acido.lattico
ADP+energia=ATP
tutto ciò avviene quando si effettua uno sforzo intenso e prolungato e
l'eccessivo accumulo di acido lattico nel muscolo può provocare il blocco quasi
completo dell'attività del muscolo stesso (crampi).
Quando, quindi, si compie uno sforzo a cui non si è abituati, a distanza di ore
o addirittura di giorni, si accusano dolori ai muscoli sollecitati dallo sforzo.
Ciò è causato dall'accumulo di acido lattico, prodotto dal processo anaerobico lattacido,.non
ancora smaltito.Per poter eliminare l'acido lattico accumulato l'unico sistema
è il riposo in modo che la circolazione sanguigna abbia la possibilità di eliminare
gradatamente questo fenomeno che ha pressocchè le stesse caratteristiche di un
avvelenamento, naturalmente temporaneo in quanto, in breve tempo, questo acido
viene ritrasformato e la situazione torna normale.
Che cosa significa allenamento
Sia che il vostro fine sia migliorare
le prestazioni in gara, sia che vogliate solo stare in piena forma psicofisica,
sia che desideriate perdere peso, o diventare campioni nel vs sport preferito,
la strada per poter raggiungere il vostro specifico scopo è l'Allenamento. Con
tale termine si intende "quell'insieme delle tecniche che consentono ad un
individuo la realizzazione massima del suo potenziale genetico, attraverso
l'apprendimento di una corretta gestualità e la razionale ripetizione di
esercitazioni mirate a modificare l'equilibrio organico individuale per
ripristinarlo ad un livello superiore". Molti di voi si chiederanno ora il
significato di questa definizione fin troppo tecnica e se da essa possiamo
trarne qualche vantaggio per la realizzazione dei nostri scopi.
a) Sicuramente, il merito o la colpa di come noi siamo oggi, lo si deve in gran
parte alle informazioni scritte nel DNA di ciascuno di noi, per cui nel nostro
codice genetico, che ereditiamo dai nostri genitori, vi sono impresse le
codifiche della ns vita, quindi la nostra altezza, la predisposizione ad essere
grassi o a contrarre le malattie, ad avere un particolare tipo di carattere,
avere il seno grosso o piccolo, le gambe corte o lunghe, eccetera.
Naturalmente, le condizioni di vita che troveremo nel corso della nostra
esistenza (specie durante i primi anni di vita, arrivando poi fino
all'adolescenza), ci aiuteranno a realizzare queste informazioni. In altre
parole, tutte le carenze che colpiscono il bambino, andranno ad incidere
nell'uomo di domani, infatti, lo scarso apporto nutrizionale, sia quantitativo
che qualitativo, le carenze affettive, gli infortuni medio gravi, la mancanza
di stimoli, la mancanza totale di movimento, non consentiranno la perfetta
"trascrizione" del DNA.
b) Altro elemento caratteristico è l'esecuzione del gesto motorio, la Tecnica.
Essa è fondamentale negli sport e gioca spesso la differenza tra scarso atleta
e campione. L'importanza è riscontrabile anche nel fitness, perché il gesto
eseguito con precisione ci aiuta a prevenire traumi, infortuni ed è garanzia di
risultato.
c) La distribuzione dei "lavori" non avviene in modo casuale, ma tiene in
considerazione delle capacità di adattamento dell'organismo. Le attività
proposte devono essere prima pensate a tavolino, ogni esercitazione deve avere
uno scopo preciso. L'insieme di queste attività andranno poi organizzate e
programmate nel tempo (Si parla allora di pianificazione, quando formuliamo una
strategia in vista di uno scopo, che poi spesso è quello di migliorarsi, di
incrementare la prestazione in gara, stabilendo obiettivi a breve a medio ed a
lungo termine e di periodizzazione, con cui intendiamo la distribuzione
cronologica dei contenuti dell'allenamento).
d) Il modo in cui noi progrediamo, è dovuto alla presenza di uno stimolo
allenante, che agisce "stressando" l'organismo. Ciò causa una momentanea
perdita di forma, a questa, se seguirà un adeguato riposo si avrà come
risultato, un miglioramento rispetto le condizioni di partenza. Perché ci sia
miglioramento, lo stimolo allenante deve essere proposto con costanza e in modo
progressivo, esso aumenta in base alle nostre capacità organiche individuali,
ciò consentirà di mantenere la sua capacità allenante e non deve avere
interruzioni prolungate per non perdere gli adattamenti raggiunti, a tal
proposito è importante alternare periodi in cui vengono somministrati stimoli
allenanti specifici a periodi di recupero attivo e di diminuzione del lavoro.
Nel caso in cui le attività proposte siano troppo "pesanti", il pericolo sarà
quello di infortunarci o nel migliore dei casi, di andare in OVERTRAINING.
L'allenamento deve avere basi scientifiche molto profonde, ma la buona riuscita
di un buon programma, dipende sia dalla osservanza di alcune regole
metodologiche sia dalla loro inosservanza.
|
Privacy |
Articolo informazione
Commentare questo articolo:Non sei registratoDevi essere registrato per commentare ISCRIVITI |
Copiare il codice nella pagina web del tuo sito. |
Copyright InfTub.com 2025