|
|
| |
IL CICLO ESTRALE
|
CICLO ESTRALE |
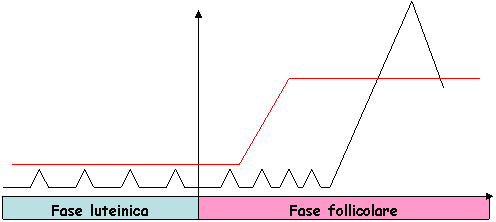
Fase passiva o luteinica |
Caratterizzata dalla presenza del CL e da alti livelli di P4 Inibizione dell'utero e dell'asse ipotalamo ipofisi |
|
Fase attiva o follicolare |
Caratterizzata dall'assenza del CL e dalla presenza di follicoli Alti livelli di 848b14i estrogeni Inizia con la regressione del CL |
Durante la fase luteinica il CL produce l'ormone P4 che ha agisce a livello periferico sull'apparato riproduttore e a livello centrale sull'asse ipotalamo-ipofisi.
A livello dell'apparato riproduttore il progesterone:
inibisce il miometrio così da avere una bassa contrattilità dell'utero. L'ormone P4 abbassa il potenziale d'azione delle cellule miometriali e blocca le giunzioni gap. In tal modo la distensione dell'utero causata dallo sviluppo del feto non viene accompagnata da contrazioni muscolari.
a livello della ghiandola mammaria stimola lo sviluppo delle strutture ghiandolari, quindi la produzione di latte
A livello dell'asse ipotalamo-ipofisi il progesterone inibisce la produzione di ormoni. In particolare, P4 inibisce la secrezione di GnRH da parte dell'ipotalamo, che lo rilascia con pulse a bassa frequenza (~ 8 ore) e ampi picchi. Questo tipo di secrezione determina una ridotta produzione di LH (fase tonica). Per quanto riguarda l'FSH questo continua ad essere secreto in modo costante, comunque le sue concentrazioni in circolo sono basse.
Questo particolare assetto endocrino si ha in presenza di un CL funzionante, quindi è utile per sostenere la gravidanza e parallelamente non è compatibile con l'attivazione della fase follicolare.
Ad un certo punto l'organismo fa una verifica per rilevare la presenza dell'embrione, quindi dell'avvenuta fecondazione. Se è avvenuta la fecondazione il CL persiste in quanto vengono prodotti ormoni che segnalano la presenza del feto. Se non c'è stata fecondazione, si attiva un meccanismo luteolitico; viene prodotta la prostaglandina PGF2α che induce la regressione del CL e blocca la produzione di progesterone.
Con la regressione del CL inizia la fase follicolare.
L'assenza del P4 determina modificazioni dell'assetto endocrino che deve essere idoneo allo sviluppo follicolare. A questo punto l'ipotalamo secerne il GnRH con pulse ogni 1-2 ore (secrezione fasica). Il GnRH stimola l'adenoipofisi a produrre LH e FSH. In questa fase l'LH viene secreto in modo fasico, cioè con pulse frequenti.
Nel momento in cui LH e FSH vengono prodotti ad alti livelli, i follicoli antrali vengono stimolati a svilupparsi.
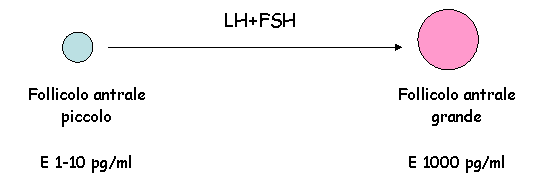
Le gonadotropine agiscono sul follicolo antrale. Si ha:
 Stimolazione
dell'attività proliferativa (mitogenica) delle cellule della granulosa e della
teca, quindi aumenta il diametro del follicolo. Le cellule della granulosa
hanno recettori per FSH mentre le cellule della teca hanno recettori per LH.
Stimolazione
dell'attività proliferativa (mitogenica) delle cellule della granulosa e della
teca, quindi aumenta il diametro del follicolo. Le cellule della granulosa
hanno recettori per FSH mentre le cellule della teca hanno recettori per LH.
Stimolazione della produzione di alti livelli di estrogeni.
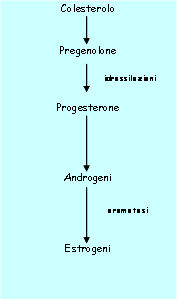
La teca riceve il colesterolo che viene convertito in androgeni. Gli androgeni passano per diffusione alla cellula della granulosa che vengono convertiti in estrogeni.
La sintesi di estrogeni è importante.
Man mano che aumentano le dimensioni del follicolo aumenta la produzione di estrogeni.
Nelle specie mono-ovulatorie e poli-ovulatorie il livello di estrogeni prodotto è simile, ciò che cambia è la dimensione dei follicoli.
I follicoli, secernendo estrogeni, segnalano il loro stadio di sviluppo all'asse ipotalamo-ipofisi. L'aumento di estrogeni ha un effetto stimolante sull'asse ipotalamo-ipofisi a produrre GnRH più frequentemente (da picchi ogni 8 ore a picchi ogni 30-10-5 minuti).
È importante in questa fase la presenza di un particolare assetto recettoriale. Ad esempio: in un animale pre-pubere l'ipotalamo non ha recettori per estrogeni mentre nelle fasi di anestro l'ipotalamo riduce i recettori per gli estrogeni e, contemporaneamente, si hanno meccanismi inibitori.
Il GnRH prodotto viene liberato ed agisce sull'adenoipofisi che produce l'LH. Il livello di LH dipende dai tempi di sintesi e degradazione. Se l'ormone viene sintetizzato con picchi frequenti e la velocità di sintesi è maggiore di quella di degradazione, si avrà un incremento notevole di LH picco LH.
È importante che la secrezione di LH sia pulsatile per avere una corretta crescita del follicolo.
Il follicolo pre-ovulatorio deve trasformarsi; le modificazioni si hanno a livello del follicolo e dell'oocita. Dal picco delle gonadotropine all'ovulazione passano diverse ore:
20-24 ore nel topo, bovino e ovino
40-44 ore nei primati, donna, maiale e cavallo
Nelle specie con un intervallo più lungo si ha un processo più complesso, probabilmente si hanno più punti di controllo. Inoltre, bisogna considerare che nelle specie con finestre temporali più brevi il macchinario necessario per il ciclo cellulare deve solo essere attivato mentre nelle specie con finestre temporali più lunghe deve essere sintetizzato.
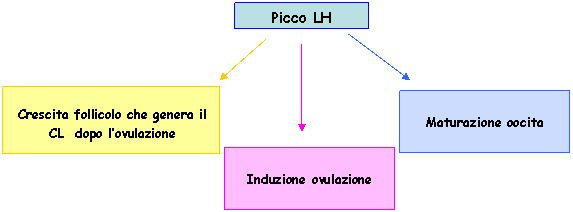
Il picco dell'LH induce diversi processi:
Il follicolo perde la capacità di produrre estrogeni. Viene persa l'aromatasi delle cellule della granulosa, quindi aumenta la concentrazione di androgeni. In un secondo momento le cellule della teca perdono la capacità di produrre androgeni, quindi si avrà solo produzione di progesterone. La produzione di progesterone inizia prima dell'ovulazione, ne consegue che quando si ha l'ovulazione ci sono le condizioni idonee alla gravidanza.
In questo cambiamento che si ha solo nel follicolo dominante è importante la modificazione dell'assetto recettoriale delle cellule follicolari. In seguito al picco dell'LH anche le cellule della granulosa presenta i recettori per l'LH (normalmente sono presenti solo sulle cellule della teca).
L'oocita matura. L'oocita termina la prima divisione e si ferma in MII, dopo essere stato ovulato potrà completare la seconda divisione solo in seguito alla fecondazione.
Il follicolo si prepara all'ovulazione. Si perde la continuità tra sierosa e membrana basale in quanto il follicolo produce enzimi proteolitici (es. collagenasi) che intaccano tali strutture. La lisi si ha in uno specifico punto del follicolo e porta alla formazione dello stigma follicolare.
In seguito all'ovulazione, il follicolo si riempie di sangue. A livello del coagulo si ha un forte processo angiogenetico e proliferazione cellulare. Il corpo luteo vero e proprio, dal punto di vista morfologico, sarà rilevabile dopo circa una settimana dall'ovulazione.
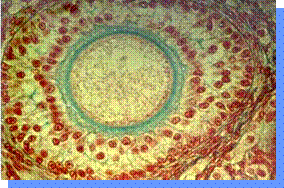 L'oocita
è circondato dalla zona pellucida (ZP) ed esternamente da cellule della
granulosa. Le cellule della granulosa
che costituiscono il cumulo ooforo sono in contatto con l'oolemma grazie a
giunzioni gap attraverso le quali possono passare sostanze nutritive e segnali
(l'oocita ha pochi recettori). La presenza di un rapporto trofico tra oocita e
cellule del cumulo è importante perché se consideriamo il rapporto
superficie/volume dell'oocita questo non è idoneo ad un adeguato assorbimento e
perché ci sono pochi recettori sull'oocita. Le cellule del cumulo, inoltre,
producono proteine che orientano lo spermatozoo verso l'oocita e a livello
della ZP ci sono proteine specifiche che permettono la reazione acrosomiale.
L'oocita
è circondato dalla zona pellucida (ZP) ed esternamente da cellule della
granulosa. Le cellule della granulosa
che costituiscono il cumulo ooforo sono in contatto con l'oolemma grazie a
giunzioni gap attraverso le quali possono passare sostanze nutritive e segnali
(l'oocita ha pochi recettori). La presenza di un rapporto trofico tra oocita e
cellule del cumulo è importante perché se consideriamo il rapporto
superficie/volume dell'oocita questo non è idoneo ad un adeguato assorbimento e
perché ci sono pochi recettori sull'oocita. Le cellule del cumulo, inoltre,
producono proteine che orientano lo spermatozoo verso l'oocita e a livello
della ZP ci sono proteine specifiche che permettono la reazione acrosomiale.
|
Privacy |
Articolo informazione
Commentare questo articolo:Non sei registratoDevi essere registrato per commentare ISCRIVITI |
Copiare il codice nella pagina web del tuo sito. |
Copyright InfTub.com 2025