|
|
| |
|
|
|
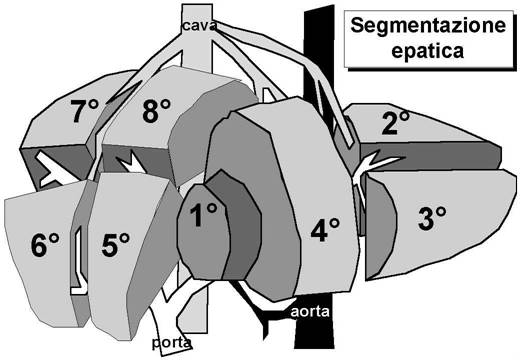
Segmentazione epatica
Il 1° segmento corrisponde al lobo caudato
situato posteriormente ed al davanti della cava. Il 2° segmento corrisponde
alla parte posteriore del lobo di sinistra. I successivi segmenti si dispongono
in senso orario rispetto ai primi (il 3° al davanti del 2°, il 4° a sinistra
del 3°, il 5° a sinistra del 4°, il 6° a sinistra del 5°, il 7° posteriormente
al 6° ed infine l'8° a destra del 7° e posteriormente al 5°).
Il 2° ed il 3° sono i segmenti mediali mentre il 6° ed il 7° sono i laterali.
Fegato
E' l'organo più voluminoso dell corpo. Una delle principali caratteristiche del fegato è la vascolarizzazione. Il sangue raggiunge quest'organo attraverso due correnti d'afflusso e lo lascia attraverso un'unica corrente di deflusso per versarsi nella vena cava inferiore.
I vasi sanguiferi che vanno al fegato sono l'arteria epatica (tronco celiaco), e la vena porta. Dal fegato defluiscono le vene epatiche. L'arteria epatica e la vena porta penetrano nel fegato in corrispondenza dell'ilo, le vene epatiche si aprono invece nella vena cava a livello della faccia post dell'organo. A livello dei sinusoidi epatici avviene mescolanza tra sangue arte e venoso: nel fegato circola sangue arterio-venoso. Il sangue venoso è drenato infine dalle vene epatiche.
La ragione di questa organizzazione vascolare è che la vena porta è il tronco venoso che conduce al fegato il sangue refluo dalla porzione sottodiaframmatica del canale alimentare (esofago addominale, stomaco, intestino tenue e crasso), e dalla milza. Q, attraverso la vena porta, il fegato riceve tutti i prodotti d'assorbi intestinale, inoltre, dal momento che nella milza avvengono fegato di eritrocateresi dovuti all'attività dei macrofagi splenici, al fegato giungono i prodotti di degradazione dell'eme (bilirubina).
Il sangue arterioso porta al fegato ossigeno e metaboliti.
Nel fegato sono presenti cellule di Kupfer (10 milioni /gr, il fegato pesa 1500 gr), che hanno posizione e funzione strategica: proseguono il lavoro dei macrofagi splenici, infatti sono in grado di produrre bilirubina a partire da frammenti di eritrociti, che possono essere arrivati al fegato attraverso la vena porta. Le cellule di Kupfer sono macrofagi residenti.
Il sangue che viene in contatto con gli epatociti è artero-venoso, ha minor ricchezza in ossigeno, esso tuttavia, è ricco di prodotti di assorbimento intestinale.
Un'altra caratteristica della vascolarizzazione è che l'arteria epatica e la vena porta, una volta penetrate nel fegato in corrispondenza dell'ilo, si dividono i 626f58g n rami che si distribuisce a grossi territori di parenchima detti zone. Ciascuna zona è caratteristiche da vascolarizzazione indipendente rispetto alle zone vicine.
Queste doti di segmentazione del fegato su base vascolare sono importanti da un punto di vista pratico, in quanto permettono di realizzare vivisezioni limitate del fegato (epatoctomie parziali).
Le zone a loro volta si suddividono in lobuli o acini, a seconda di come si considera la costituzione del fegato.
Per le complesse funzioni, il fegato può essere considerarsi non solo come un ghiandola esocrina, in quanto elabora la bile che riversa nel duodeno per mezzo dei condotti biliari, ma anche come una particolare ghiandola endocrina, non perché secerne ormoni ma perché riversa direttamente nel sangue numerosi elaborati (proteine plasmatiche, glucoso, lipoproteine).
Il parenchima epatico è costituito dagli epatociti. Gli epatociti sono cellule polarizzate, in quanto in essi è possibile individuare un polo vascolare, in corrispondenza del quale l'epatocita opera selettivamente scambi con il sangue, ed un polo biliare, in corrispondenza del quale avviene la secrezione della bile.
Negli epatociti esiste inoltre una precisa compartimetalizzazione, cioè una precisa distribuzione degli organelli in base alle funzioni. Il RER opera per la produzione delle proteine plasmatiche (+ di 80) riversate in circolo, il REL opera per la sintesi del colesterolo e la degradazione dei farmaci liposolubili (funzione detossificante).
FUNZIONI
1) biligenesi. Il fegato elabora la bile che, attraverso le vie biliari, viene immessa nel duodeno, dove svolge importanti funzioni, in particolare ai fini della digestione dei grassi. La bile è anche vettore dei prodotti che devono essere eliminati e di prodotti fondamentali. Essa è arricchita di IgA, e l'80% di IgA presenti nel tenue arriva con la bile.
2) Il fegato è intercalato tra il circolo portale e quello della vena cava inferiore, riceve il sangue refluo dalla milza, stomaco, tenue e dalla maggior parte del crasso ed opera sui metaboliti assorbiti a livello intestinale, fungendo come organo di deposito per alcuni materiali (es. glicogeno), che hanno un importante ruolo nel mantenimento di parametri ematochimici (es. glicemia). In questo senso, il fegato può es considerato come un'organo essenziale per mantenere l'omeostasi ematica.
3) funzioni metaboliche.
a) metabolismo dei glucidi (gluconeogenesi, glicogenosintesi e glicolisi, che sono alla base della funzione glicostatica del fegato e quindi della regolazione della glicemia).
b) metabolismo dei lipidi (sintesi dei trigliceridi, acidi grassi e lipoproteine, formazione dei corpi chetonici, sintesi, degradazione, esterificazione ed escrezione del colesterolo)
c) metabolismo delle proteine (sintesi delle proteine plasmatiche, del fibrinogeno, della protrombina, di quasi tutti i fattori plasmatici della coagulazione, catabolismo delle proteine e dei relativi aa).
L'albumina (4/5 g/100mL) è responsabile del mantenimento della pressione oncotica dei protidi plasmatici. La diminuzione dell'albumina plasmatica può comportare riduzione della pressione oncotica e pertanto alterazione degli scambi idrici tra sangue e tessuti, causando l'abnorme passaggio di liquidi nei tessuti, arrivando alla formazione dell'edema.
d) metabolismo dei composti azotati non proteolitici (sintesi urea, ac. urico, purine pirimidine, glutatione)
e) metabolismo delle vitamine (fosforillazione della tiamina, piridossale, riboflavina, formazione coenzima A dall'acido pantoteico e dal coenzima B12 della vitamina B12)
4) funzione detossificante e trasformante. Molte sostanze tossiche, o farmaci (es. barbiturici), possono essere eliminati da enzimi che, nel caso dei barbiturici, sono localizzati a livello del REL. Infatti, esaminando il fegato di un soggetti che ha fatto uso di barbiturici, si può constatare una notevole estensione del REL negli epatociti.
La funzione trasformante si realizza attraverso reazioni di ossidazione (perossisomi),... Da notare che il fegato può, idrossilando composti del tutto innocui, creare sost cancerogene.
5) funzione di coniugazione (con acido glucuranico). Tipicamente la bilirubina proveniente dai macro splenici e dalle cellule di Kupfer deve esse glucurono coniugata per finire nella bile. Avremo così una bilirubina che non è ancora passata per il fegato, ed una bilirubina diretta, glucurono coniugata.
6) mantenimento omeostasi. Esistono parametri emolitici per i quali è necessaria la normale funzione del fegato.
Si è visto come il fegato può essere suddiviso in zone, che presentano vascolarizzazione indipendente, l'ulteriore suddivisione in lobuli non presenta le stesse caratteristiche, in quanto i lobuli presentano una vascolarizzazione multipla.
I lobuli sono ben evidenti sopratutto nelle specie animali in cui il fegato è provvisto di una abbondante trama di connettivo, essi vengono comunemente considerati come le unità strutturali del fegato (lobuli classici). Ogni lobulo risulta costituito da lamine cellulari che delimitano un sistema labirintico in cui è contenuta una reta capillare di vasi sanguiferi a decorso tortuoso, i sinusoidi.
Nel lobulo classico le lamine cellulari ed i sinusoidi hanno disposizione radiale: dalla periferia del lobulo convergono verso il centro.
L'asse del lobulo epatico è occupato da un vaso venoso a parete sottile, la vena centrolobulare, in cui sboccano tutti i sinusoidi contenuti nel lobulo epatico.
La parete della vena centrolobulare appare perciò discontinua. Le zone dove lobuli adiacenti vengono tra loro in contatto sono indicate come spazi portali, in cui decorrono i rami lobulari dell'arteria epatica, della vena porta ed i canalicoli biliari. Queste tre formazioni costituiscono la triade portale.
Il sangue che circola nelle ramificazioni dell'arteria epatica e della vena porta viene convogliato dalla peri del lobulo epatico nella rete di sinusoidi.
Sino a livello della triade il sangue arterioso e quello venoso circolano indipendentemente e le tre formazioni decorrono all'interno di tralci connettivali molto evidenti. I sinusoidi si portano alla vena centrolobulare, che percorre assialmente il lobulo, alla base dei lobuli, le vene centrolobulari confluiscono nelle vene sottolobulari (circolo venoso puro), tributaria a loro volta delle vene epatiche, che sboccano nella vena cava inferiore. Le ramificazioni dell'arteria epatica e della vena porta confluiscono nello stesso sistema di drenaggio quando si immergono nei sinusoidi. Questi ultimi sono riccamente anastomizzati per cui il sangue che circola nei sinusoidi è arterovenoso.
La bile elaborata dagli epatociti si riversa dentro spazi intercellulari scavati tra le pareti di epatociti contigui (capillari biliari), quindi viene convogliata in condotti provvisti di parete propria presenti negli spazi portali (canalicoli biliari).
Come nei seni venosi della polpa rossa, anche nei sinusoidi epatici la velocità del sangue è modesta: nel passaggio da triade portale a rete sinusoidale si ha una grassa caduta di pressione e rallentamento del flusso, ciò permette agli epatociti di restare a lungo contatto con il sangue.
MODELLI DI ORGANIZZAZIONE DEL PARENCHIMA EPATICO
Il lobulo epatico (classico) è caratterizzato dagli spazi portali, con triade portale (ramo lobulare di arteria epatica e vena porta, canalicolo biliare).
Questo modello non è del tutto soddisfacente e presenta alcuni limiti.
Nel caso per es di tumore della testa del pancreas, o do ostruzione delle vie biliari, si verifica un ristagno di bile, e la formazioni di lesioni triangolari, con un canalicolo biliare al centro e vene centrolobulari ai vertici.
Il secondo modello (del lobulo portale) è caratterizzato da un territorio di parenchima il cui centro è occupato da un canale biliare, questo è contenuto nello spazio portale a raccogliere la bile secreta dall'area circostante di parenchima epatico. I limiti del lobulo epatico si possono ottenere congiungendo con una linea immaginaria tre vene centrolobulari. Questo modello di organizzazione del parenchima epatico pone l'accento sulla funzione esocrina del fegato. In esso il sangue scorre in direzione centrifuga (dallo spazio portale alla vena centrolobulare), mentre la bile ha decorso centripeto (dalla periferia all spazio portale).
Anche questo modello non è del tutto soddisfacente, in quanto non è in grado di spiegare una delle principali patologie che colpiscono il fegato, la cirrosi epatica.
Nel caso di questa malattia non solo viene distrutto il parenchima epatico, ma il connettivo va a strozzare i vasi arteriosi e venosi, impedendo la vascolarizzazione di queste zone. La rigenerazione del parenchima epatico (vedi oltre), si realizza a partire da rami preterminali e terminali dell'arteria epatica e della vena porta, a seconda che lo strozzamento, e quindi la degenerazione sia avvenuta a livello preterminale o terminale, la rigenerazione inizierà a monte della strozzatura. Avremo allora la formazione di acini, che saranno semplici se siamo a livello terminale, complessi a livello preterminale.
Questi modelli di organizzazione del parenchima epatico sono l'acino semplice e complesso, cioè territori che presentano al centro rami preterminali o terminali della vena. porta, in quanto è da questi rami che ha inizio il processo di rigenerazione.
Un ultimo concetto riguarda la polarità degli epatociti: la superficie degli epatociti rivolta verso i sinusoidi è provvista di microvillo (aumento della superficie assorbente), in corrispondenza del polo biliare, la mb dell'epa presenta una depressione foggiata a doccia. Questa depressione si guistappone ad un'analoga presente sulla parete dell'epatocito adiacente, si delimita in questo modo il capillare biliare, sprovvisto di parete propria.
Superfici relativamente pianeggianti permettono un'esatta giustapposizione con gli epatociti circostanti.
ANATOMIA
Il polo vascolare è la superficie dell'epatocita in rapporto con i sinusoidi, la membrana dell'epatocita è sollevata in numerosi microvilli.
I sinusoidi sono capillari sanguigni la cui parete è costituita da cellule endoteliali organizzate in un dispositivo discontinuo, sono riccamente anastomizzati.
Convogliano il sangue dai rami lobulari dell'arteria epatica e della vena porta, collocate nello spazio portale, alla periferia del lobulo, verso la vena controlobulare che percorre assialmente il lobulo.
La parete dei sinusoidi è costituita da cellule endoteliali appiattite, che sporgono nel lume solo con quella porzione in cui è contenuto il nucleo. L'endotelio è discontinuo per la presenza di pori e fenestrature.
Tra l'epatocita e la parete del sinusoide si trova lo spazio di Disse. Attraverso le fenestrature dell'endotelio, il plasma filtra nello spazio di Disse e si pone in rapporto con l'epatocita.
Lo spazio di Disse si estende anche tra due epatociti contigui, e permette un contatto prolungato del plasma con la parete dell'epatocita, sollevata in numerosi microvilli.
Il contatto deve esse lento e prolungato, in quanto a questo livello avvengono numerosi scambi tra epatocita e plasma: acidi grassi vengono assorbiti, lipoproteine e proteine plasmatiche vengono rilasciate.
La struttura della parete dei sinusoidi non è così semplice: i sinusoidi epatici non presentano una membrana basale così come i sinusoidi splenici. L'assenza della membrana rende ragione della permeabilità di questi vasi che nella milza permettono il riassorbimento del plasma, e nel fegato permettono il passaggio del plasma nello spazio di Disse, quindi il ritorno dello stesso plasma "richiamato" dal flusso sanguigno.
Nello spazio di Disse si trovano fibre reticolari, che formano un reticolo di supporto per la parete del sinusoide.
Nei sinusoidi, sporgenti nel lume, si trovano elementi con attività fagocitica attiva, le cellule di Kupfer. Si trovano nel contesto delle cellule endoteliali, ma non sono in linea, si trovano sul versante interno dell'endotelio, appoggiate sulle cellule endoteliali, appartengono alla famiglia dei monociti macrofagi. Sono elementi residenti e presentano forma irregolare, a cavallo del sinusoidi. Aderiscono alla superficie interna dell'endotelio e vanno a popolare la superficie interna dei sinusoidi.
Le cellule di Kupfer si spostano all'interno dei sinusoidi: è necessario un continuo avvicendamento.
Pur essendo macrofagi residenti possono staccarsi dall'endotelio e tornare ad essere monociti circolanti. Esiste, dunque, un continuo ricambio di questo tipo di cellule all'interno del fegato. Completano l'azione emocateretica dei macrofagi splenici.
Queste cellule, inoltre, hanno l'importante ruolo di cellule presentanti l'antigene (come le cellule interdigitate e cellule dendritiche follicolari).
Una piccola parte di queste cellule, che processano antigeni i quali vengono riespressi nel contesto di MHC II, è in grado di raggiungere i linfonodi dell'ilo epatico. Nel parechima epatico, infatti, non vi sono vasi linfatici (presenti sino agli spazi portali),e solo cellule di Kupfer situate alla periferia dei lobuli possono terminare nella zona dell'ilo.
I linfonodi di questa zona presentano una paracortex particolarmente sviluppata (zona T dipendente), e questo è dovuto al continuo apporto di antigeni (espressi nel contesto di MHC II) portati da cellule di Kupfer.
Oltre alle fibre reticolare, nello spazio di Disse si trovano anche cellule che hanno la caratteristica proprietà di accumulare nel cito lipidi. Si pensa che queste cellule siano un serbatoio per le sost liposolubili come le vitamine A e K (importante nella coagulazione). Questi elementi sono netti cellule di ITO o fat storng cells.
Le cellule di ITO differiscono dalle cellule di Kupfer per sede e funzioni: si trovano all'esterno e non all'interno dell'endotelio che delimita i sinusoidi.
Sino ad ora abbiamo considerato il polo vascolare degli epatociti, consideriamo ora quelle zone in cui viene dismessa la bile: è possibile visualizzare queste zone trattando le cellule con metodi specifici per la dimostrazione di enzimi di membrana a funzione ATPasica, necessari al meccanismo delle pompe ioniche. Si ottiene così una mappa molto articolata che evidenzia quelle formazioni canaliformi costituite dalla giustapposizione,e di epatociti adiacenti, che presentano sulle membrane solchi foggiati a doccia, i capillari biliari.
La bile viene riversata nel capillari biliari, alla periferia dell'lobulo si trovano territori che rappresentano il punto di passaggio tra capillari biliari, privi di parete propria, e canalicoli biliari, provvisti di parete propria e situati negli spazi portali.
Questo segmento di transito è detto colangiolo.
I capillari biliari non presentano parete propria, ma in corrispondenza di questi la membrana dell'epatocita si estroflette in microvilli, ed inoltre presentano enzimi di membrana ad attività ATPasica.
Pur non possedendo una parete propria, i capillari biliari devono esse "chiusi" in qualche modo per impedire che la bilirubine già passata negli epatociti, quindi coniugata con acido glucuronico, filtri negli spazi di Disse e torni nel circolo ematico.
Questo fenomeno si verifica in patologie come i calcoli delle vie biliari, che causano una elevata pressione a livello del capillari, I quindi una fuoriuscita di bilirubina diretta. i sintomi di questa patologia sono dati da un ittero molto evidente. L'ittero può perciò esse causato anche dalla presenza nel sangue di bilirubina indiretta (non coniugata).
In questo caso l'ittero è dovuto a patologie molto diverse, per es una malattia emolitica acuta che causa un elevato afflusso di bilirubina al fegato ed il mancato riassorbimento da parte degli epatociti.
In condizioni di normalità, i capillari biliari sono strutture perfettamente chiuse, grazie ai classici dispositivi di giunzione (presenti anche a livello degli enterociti), ovvero la triade sona occludente, zona aderente, desmosoma.
All'interno degli epatociti, i lisosomi sono localizzati in corrispondenza dei capillari biliari, e questo sempre per il concetto di polarizzazione e compartimentazione, definito per gli epatociti: i lisosomi infatti sono fondamentali, con la loro attività idrolasica, in diverse tappe delle biligenesi.
In modo analogo, il reticolo endoplasmatico e l'apparato di Golgi sono localizzati in corrispondenza del polo vascolare dell'epatocita.
Un'altra caratteristica del polo biliare degli epatociti è la presenza di numerosi filamenti di cheratina. Questi costituiscono il supporto dei capillari biliari, e fanno in modo che i capillari siano una struttura rigida: questo è importante in quanto se i capillari avessero una struttura flessibile, un lieve aumento di pressione causerebbe la distensione delle pareti del capillare, e l'indebolimento dei dispositivi di giunzione che devono garantire una perfetta "chiusura" dei capillari, i quali non possiedono una parete propria.
Passiamo ora ai colangioli. Si tratta di condotti che cominciano ad avere una parete propria, costituita da cellule epiteliali e dalla superficie degli epatociti. In pratica i colangioli sono costituiti da una singola cellula che con i suoi prolungamenti cito va a costituire una struttura vasale.
Queste cellule sono molto importanti per due motivi: esiste una situazione patologica per cui, durante la morfogenesi delle vie biliari, non si formano i colangioli. L'agenesia del colangiolo determina il mancato smistamento della bile, con conseguente morte, a meno di un rapido trapianto di fegato. Inoltre le cellule dei colangioli sono elementi staminali in gradi di differenziarsi in epatociti: da essi dipendono i fenomeni di rigenerazione, dovuti a capacità di differenziamento da parte delle cellule dei colangioli.
Lasciati i colangioli si entra nei condotti biliari che confluiscono nei due dotti epatici per poi arrivare alle vie biliari extraepatiche.
All'interno del fegato, i canalicoli biliari non presentano modificazioni rilevanti, così come non subisce modificazione la bile, almeno fino a che non arriva alla colecisti.
Le vie biliari extraepatiche sono rappresentate dal dotto epatico com, che in seguito riceve il dotto cistico, per andare a formare il dotto coledoco. Questo passa dietro la prima porzione del duodeno e la testa del pancreas, e va a sboccare a livello della faccia mediale della seconda porzione del duodeno.
A livello della papilla duodenale sono presenti formazioni sfinteriche, che permettono la dismissione di bile solo nel caso di presenza di grassi nel duodeno, ed ulteriori sfinteri che impediscono il passaggio della bile nel condotto pancreatico principale quindi nel pancreas, con la conseguente attivazione degli enzimi proteolitici che distruggerebbero il pancreas (pancreatiti acute).
La parete di questi condotti è costituita da epitelio di rivestimento e da lamina propria, con infiltrazione lifocitaria (MALT), ed alcune ghiandole. Esternamente alla mucosa si trova una tonaca fibromuscolare, la componente muscolare è necessaria alla progressione della bile nei dotti.
Colecisti
La parete di quest'organo presenta alcune caratteristiche dovute al fatto che in esso la bile ristagna, subendo modificazioni sostanziali.
La mucosa è sollevata in pieghe ramificate, alla cui formazione prende parte anche la lamina propria. L'epitelio di rivestimento è batiprismatico semplice.
La mucosa si mette in rapporto con la tonaca fibromuscolare senza interposizione di una sottomucosa.
La tonaca fibromuscolare è costituita da fasci di fibrocellule muscolari lisce i,ntrecciate (si determina la spremitura dell'organo), e da fibre collagene.
In fine è presente una tonaca avventizia sulla faccia superiore, una sierosa peritoneale sulla faccia inferiore.
Nel contesto della mucosa non sono presenti ghiandole, l'epitelio di rivestimento è caratterizzato da un'orletto striato breve e sottile, formato da corti microvilli.
Le cellule epiteliali sono saldamente unite tra loro a livello delle parti apicali mediante complessi di giunzione (zonula aderente, occludente, desmosoma).
A queste aree, che devono esse considerate impermeabili, fanno seguito, verso il polo basale, la membrana caratterizzata da un profilo irregolare per la presenta di interdigitazioni che delimitano uno spazio canalicolare intercellulare esteso sino alla membrana basale su cui le cellule poggiano.
La colecisti non funziona solo come un serbatoio, ma serve anche a concentrare la bile assorbendo parte dell'acqua in essa contenuta.
Mentre nella fasi di riposo il sistema dei canalicoli intercellulari, che si delimita tra le cellule dell'epitelio di rivestimento appare ridotto ad una sottile fessura, nella fase di concentrazione della bile (trasporto attivo di Na+ e passivo di acqua) gli spazi canalicolari intercellulari risultano dilatati.
Nel plasmalemma delle superfici laterali delle cellule epiteliali è stata dimostrata attività ATPasica. Il trasporto attivo degli elettroliti, l'assorbimento di acqua ha luogo a seguito del trasporto attivo di Na+ negli spazi intercellulari. Si stabilisce un gradiente osmotico che richiama acqua dal lume della colecisti.
In seguito avviene il drenaggio dei fluidi: spazi intercellulari, membrana basale, capillari sanguigni, tonaca propria.
|
Privacy |
Articolo informazione
Commentare questo articolo:Non sei registratoDevi essere registrato per commentare ISCRIVITI |
Copiare il codice nella pagina web del tuo sito. |
Copyright InfTub.com 2025