|
|
| |
GENETICA MENDELIANA
Sebbene l'ereditarietà biologica sia
stata oggetto di interesse e stupore sin dagli inizi della storia umana, solo
recentemente l'uomo ha iniziato a capire il suo funzionamento; in effetti lo
studio scientifico dell'ereditarietà, noto come genetica, non iniziò di fatto
prima della seconda metà del 1800 con il monaco Gregor Johann Mendel
(1822-1884). Il suo lavoro, effettuato nel giardino di un tranquillo monastero
agostiniano nell'attuale Brno, segnò l'inizio della genetica moderna.
Il maggior contributo di Mendel fu l'aver dimostrato che i caratteri ereditari
sono trasmessi come unità che vengono distribuiti singolarmente a ogni
generazione. Queste unità distinte, che vengono da Mendel Elemente,
furono in seguito chiamate geni.
La soluzione che dette Mendel al problema dell'eredità sembra oggi tanto
semplice che sorprende che nessuno nel 1865 avesse compreso la sua accurata e
ragionata analisi. Quel lavoro, che segnò l'inizio della biologia quantitativa,
rimane ancora come un modello di brillante procedura sperimentale.
Mendel osservò l'andamento numerico di alcune caratteristiche di tre
generazioni di piante di pisello ed, in seguito, analizzò matematicamente i
risultati ottenuti. E' proprio questa l'innovazione di Mendel: l'idea che un
problema biologico potesse essere studiato quantitativamente era del tutto
nuova; poi, analizzando i risultati ottenuti, ipotizzò due leggi, note come"
legge della segregazione" e "legge della segregazione indipendente" , che
diventarono le prime importanti leggi di genetica, e quindi diedero di fatto
origine alla genetica classica.
La scelta di Mendel di utilizzare la pianta di pisello per i suoi esperimenti
non era certo originale. Tuttavia egli riuscì a fo 949j92j rmulare i princìpi
fondamentali dell'ereditarietà, dove altri avevano fallito, grazie al suo
approccio metodologico. Innanzitutto egli verificò un'ipotesi molto specifica
in una serie di esperimenti logici. Pianificò i suoi esperimenti con cura ed
intelligenza, scegliendo di studiare solamente differenze ereditarie nette e
scartando le caratteristiche che potevano apparire nella prole in modo incerto.
In secondo luogo, Mendel studiò i discendenti non solo della prima generazione
e della seconda, ma anche delle generazioni successive. Infine, in terzo luogo,
analizzò i suoi dati in modo tale da rendere la loro valutazione semplice ed
oggettiva. Gli esperimenti stessi furono descritti così chiaramente che
poterono essere ripetuti e controllati da altri scienziati, cosa che in effetti
poi avvenne.
INCROCI SPERIMENTALI
Mendel selezionò sette caratteri che mostravano, nelle diverse varietà di
piante di pisello, due forme nettamente differenti. Una varietà, per esempio,
produceva sempre semi gialli, mentre un'altra sempre semi verdi. In seguito,
Mendel esegui incroci sperimentali asportando le antere di un fiore contenenti
il polline e cospargendo gli stigmi con il polline di un fiore di un altra
varietà. Poi Mendel permise ai fiori di pisello di autoimpollinarsi, quindi di
dare origine ad un'altra generazione da analizzare.
LEGGE DELLA SEGREGAZIONE
Mendel mise a confronto le
generazioni da lui analizzate ed osservò che nella prima generazione F , cioè «prima generazione filiale», tutti i figli mostravano solamente
uno dei caratteri presenti nei genitori; l'altro carattere era completamente
scomparso.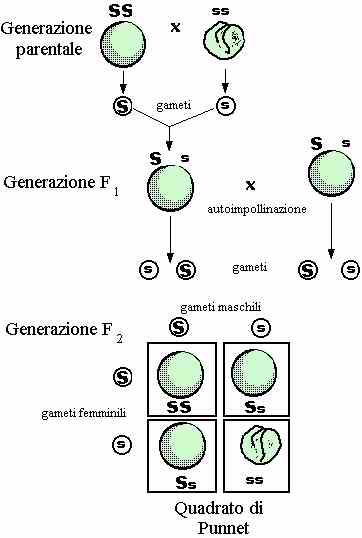
Le caratteristiche che apparivano nella generazione F1 furono chiamate da
Mendel dominanti. Però a questo punto sorse spontanea una domanda: che
cosa era successo al carattere antagonista? Il quesito fu risolto dall'analisi
della «seconda generazione filiale» o F , in
cui riapparivano i caratteri scomparsi nella generazione precedente. Queste
caratteristiche, presenti nella generazione parentale (P) e ricomparse
nella F , dovevano in qualche modo essere presenti anche nella generazione F ,
sebbene non evidenti. Mendel chiamò questi caratteri recessivi. La F
quindi era composta da caratteri sia dominanti che recessivi, però legati dal
rapporto 3:1. Mendel intuì che la comparsa dei caratteri antagonisti e le loro
proporzioni costanti nella F potevano essere spiegate
ammettendo che le caratteristiche fossero determinate da fattori separati.
Questi fattori, riteneva Mendel, dovevano trovarsi nelle piante F
in coppie: un componente di ogni coppia era ereditato dal padre e l'altro dalla
madre. Questa, nota anche come prima legge di Mendel, è la legge della
segregazione.
Quindi la F , dovendo avere entrambi i caratteri, può essere scritta come Yy,
di conseguenza chiamarla eterozigote; però c'è da ricordare che un
organismo eterozigote manifesta nel suo fenotipo (aspetto esteriore)
solo l'allele (carattere) dominante.
Mentre la P è formata da organismi yy e YY, cioè da linee pure,
chiamati anche omozigoti.
Chiarito il significato di tali parole possiamo cercare una spiegazione del
rapporto 3:1. Uno dei modi più semplici è il quadrato di Punnet, dal
nome del genetista inglese che per primo o utilizzò per l'analisi dei caratteri
determinati geneticamente. Il quadrato di Punnet utilizza le leggi della
probabilità.
LEGGE DELL'ASSORTIMENTO INDIPENDENTE
In una seconda serie di esperimenti
Mendel prese in considerazione degli incroci tra piante di piselli che
differivano per due caratteri: un genitore produceva semi lisci e gialli e
l'altro rugosi e verdi. I caratteri liscio e giallo sono dominanti, mentre
rugoso e verde sono recessivi.
Mendel, seguì il medesimo procedimento che aveva usato con il primo
esperimento, ed ottenne risultati simili, ma significativi. La F
era composta per intero da componenti genotipicamente eterozigoti (quindi
fenotipicamente con semi lisci e gialli), mentre fra i componenti della F
Mendel notò un rapporto in media di 9:3:3:1. Su 16 combinazioni, 9 sono gli
individui che presentano i due caratteri dominanti, 1 è l'individuo con i due
caratteri recessivi, 3 e 3 sono gli individui con le due combinazioni
alternative di caratteri dominanti e recessivi.
In base a questi risultati, Mendel formulò la seconda legge, o dell'assortimento
indipendente, che afferma: "...quando si formano i gameti, gli alleli
di un gene si separano indipendentemente dagli alleli di un'altro gene".
GENETICA CLASSICA
Nei decenni che seguirono la riscoperta, nel
1900, degli studi di Mendel, in genetica vennero condotte numerosissime
ricerche anche grazie alle quali i biologi riuscirono a confermare e ampliare
le osservazioni di Mendel. I metodi e i dati ottenuti in questo periodo, durato
circa 50 anni costituiscono il nucleo centrale di quel ramo della biologia che
noi chiamiamo genetica classica.
La genetica classica si occupò prevalentemente di ampliare e correggere
concetti sviluppati dalle teorie di Mendel
MUTAZIONI
Nel 1902 un botanico olandese, Hugo de Vries, notò che a volte in una pianta da
lui analizzata si presentava un fenotipo del tutto nuovo, de Vries ipotizzò che
un carattere nuovo comparisse in seguito a improvvisi cambiamenti avvenuti nei
geni, e chiamo questi cambiamenti mutazioni e gli organismi con tali
mutazioni furono detti mutanti.
INTERAZIONI
INTERAZIONI ALLELICHE
Dominanza incompleta e codominanza
A mano a mano che gli studi di genetica procedevano, divenne chiaro che le
caratteristiche dominanti e recessive non sono sempre così nette come i sette
caratteri studiati da Mendel nella pianta di pisello.
Alcune caratteristiche sembrano mescolarsi: per esempio, incrociando una pianta
di bocca di leone con fiori rossi con un'altra con fiori bianchi si producono
eterozigoti di colore rosa. Questo fenomeno, in cui il fenotipo
dell'eterozigote è intermedio tra quelli dei due omozigoti, è detto dominanza
incompleta. Però in altri casi gli alleli possono agire da codominanti,
con eterozigoti che esprimono contemporaneamente entrambi i fenotipi omozigoti.
Un esempio familiare è il sangue 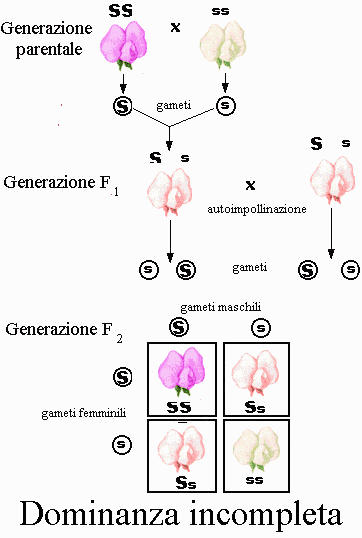 umano di tipo AB, in cui i globuli rossi mostrano le caratteristiche
relative sia al tipo A sia al tipo B.
umano di tipo AB, in cui i globuli rossi mostrano le caratteristiche
relative sia al tipo A sia al tipo B.
Alleli multipli
Ogni organismo diploide (cioè avente materiale genetico di entrambi i genitori)
può avere soltanto due alleli per ogni gene, ma in una popolazione di organismi
possono essere presenti più di due forme di un gene; si hanno perciò alleli
multipli che derivano da differenti mutazioni dello stesso gene. Negli
esseri umani i quattro principali gruppi sanguigni (A, B, AB e 0) sono
determinati da un gene che ha tre alleli (A, B e 0).
INTERAZIONI GENICHE
Oltre alle interazioni che avvengono tra alleli della stesso gene, ci sono
anche le interazioni tra alleli di geni differenti. In realtà, la maggior parte
delle caratteristiche, sia strutturali sia chimiche, che costituisce il
fenotipo di un individuo, è il risultato dell'interazioni tra due o più geni
distinti. Queste interazioni possono essere di varia natura.
Talvolta, quando un carattere è influenzato da due o più geni differenti, può
apparire un fenotipo del tutto nuovo.
In altri casi, invece, l'interazione genica non produce alcun nuovo fenotipo,
ma un gene può interferire con un altro mascherandone gli effetti. Questo tipo
di interazione è detta epistasi.
Però alcuni caratteri, come le dimensioni o la statura, la forma, il peso, il
colore, il tasso metabolico e il comportamento, non sono il risultato di
interazioni tra uno, due o più geni; sono invece il risultato complessivo degli
effetti combinati di molti geni. Questo fenomeno è detto eredità poligenica
ESISTENZA CONCRETA DEL GENE
L'ipotesi che i geni fossero posti
sui cromosomi venne definitamente confermata dagli studi effettuati sul
moscerino della frutta Drosophila, che è stato utilizzato in una grande
varietà di studi di genetica. Esso possiede quattro coppie di cromosomi; tre di
queste copie (gli autosomi) sono strutturalmente uguali per entrambi i
sessi, ma la quarta coppia, i cromosomi sessuali, è diversa. Nei
moscerini della frutta, come molte altre specie (inclusa la specie umana), i
due cromosomi sessuali sono XX nelle femmine e XY nei maschi.
Durante la meiosi i cromosomi sessuali, come pure gli autosomi, segregano (si
separano). Ogni cellula uovo riceve un cromosoma X, ma metà degli spermatozoi
riceve cromosomi X e metà Y. Perciò, è lo spermatozoo che determina il sesso dell'embrione
nelle specie con i maschi XY.
Agli inizi del XX secolo T.H. Morgan dimostrò con gli esperimenti di
incrocio sul moscerino della frutta che certi caratteri sono legati al sesso,
cioè che i loro geni sono portati sui cromosomi sessuali. Poiché il cromosoma X
porta dei geni che non sono presenti sul cromosoma Y, un singolo allele
recessivo sul cromosoma X del maschio dà luogo ad un fenomeno recessivo dal
momento che non è presente alcun altro allele. Al contrario, una femmina
eterozigote per un carattere legato al sesso mostrerà caratteristiche
dominanti.
Nell'uomo, tra i caratteri legati al sesso ci sono il daltonismo e l'emofilia.
GENETICA MOLECOLARE
I cromosomi, come tutte
le altre parti di una cellula vivente, sono formati da atomi disposti in
molecole. Alcuni scienziati, tra cui personalità eminenti nel campo della
genetica, pensavano che sarebbe stato impossibile capire la complessità dei
meccanismi ereditari sulla base della struttura di sostanze chimiche
«inanimate». Altri ritenevano che, una volta chiarita la struttura
chimica dei cromosomi, sarebbe stato possibile capire il loro ruolo come
portatori delle informazioni genetiche; questa intuizione segnò l'inizio di un
fruttuoso campo di ricerche, la genetica molecolare.
Le prime analisi chimiche del materiale ereditario rivelarono che il cromosoma
eucariote è costituito da acido deossiribonucleico (DNA) e da
proteine, sostanze presenti più o meno in uguale quantità; perciò entrambe
avrebbero potuto avere il ruolo di materiale genetico.
Le proteine sembravano essere la soluzione più probabile a causa della loro
maggiore complessità chimica; le proteine infatti sono polimeri di amminoacidi
di cui nelle cellule, se ne conoscono 20 diversi tipi. Il DNA, invece, è un
polimero formato solamente da quattro differenti tipi di nucleotidi.
LA NATURA DEL DNA
Il DNA era stato isolato per la prima volta dal medico tedesco Friedrick
Miescher nel 1869, nello stesso importante decennio in cui Darwin
pubblicava L'Origine delle Specie e Mendel comunicava i suoi risultati
alla Società di Storia Naturale di Brùnn. La sostanza isolata da Miescher
era bianca, zuccherina, leggermente acida e conteneva fosforo. Poiché era
stata trovata soltanto nei nuclei delle cellule, venne chiamata acido
nucleico. Tale nome fu in seguito modificato in acido
deossiribonucleico (DNA) per distinguere questa sostanza da una
simile, l'acido ribonucleico (RNA).
Ogni nucleotide è formato da una base azotata, dallo zucchero deossiribosio e
da una base azotate e un gruppo fosfato. Vi sono due tipi di basi azotate: le purine,
che presentano una struttura a due anelli e le pirimidine che hanno un
solo anello. Nel DNA vi sono due tipi di purine, l'adenina (A) e
la guanina (G) e due tipi di pirimidine, la citosina (C)
e la timina (T). Così il DNA è costituito da quattro tipi di
nucleotidi che differiscono soltanto per tipo di purine o di pirimidine
contenenti azoto.
Gli esperimenti sui
batteriofagi
Nel 1940 ebbe inizio una serie di esperimenti fondamentali che utilizzavano un
altro «materiale adatto», destinato a diventare tanto importante nella ricerca
quanto la pianta di pisello e il moscerino della frutta. Tale materiale
era un gruppo di virus che attaccano i batteri e sono pertanto detti batteriofagi.
(«mangiatori di batteri»). I batteriofagi scelti inizialmente per questi
studi furono quelli che attaccano Escherichia coli uno tra i più comuni
batteri dell'intestino umano.
L'analisi clinica dei batteriofagi rivelò che essi sono costituiti quasi esclusivamente
da DNA e proteine, le due sostanze che negli anni '40 erano i principali
concorrenti al ruolo di materiale genetico.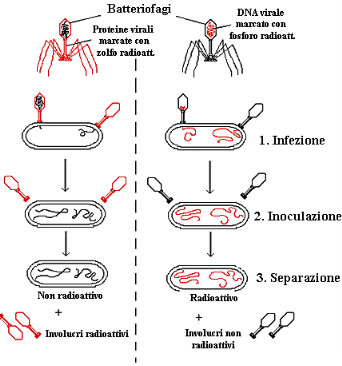 La questione su quale dei due tipi di molecola contenga i geni
virali, cioè il materiale ereditario che dirige la sintesi di nuovi virus
all'interno della cellula batterica, fu risolta nel 1952 da Alfred D. Hershey
e da Martha Chase. Osservando la figura tenete presente che, mentre le
proteine contengono zolfo (negli amminoacidi metionina e cisteina), ma non
fosforo, il DNA contiene fosforo ma non zolfo. Grazie a questi
esperimenti è stato possibile dimostrare che solamente il DNA dei batteriofagi
era coinvolto nel processo di duplicazione e che le proteine non potevano
costituire il materiale genetico.
La questione su quale dei due tipi di molecola contenga i geni
virali, cioè il materiale ereditario che dirige la sintesi di nuovi virus
all'interno della cellula batterica, fu risolta nel 1952 da Alfred D. Hershey
e da Martha Chase. Osservando la figura tenete presente che, mentre le
proteine contengono zolfo (negli amminoacidi metionina e cisteina), ma non
fosforo, il DNA contiene fosforo ma non zolfo. Grazie a questi
esperimenti è stato possibile dimostrare che solamente il DNA dei batteriofagi
era coinvolto nel processo di duplicazione e che le proteine non potevano
costituire il materiale genetico.
MODELLO WATSON-CRICK
Nel 1953 lo scienziato americano James Watson ed il fisico
francese Francis Crick proposero un modello di struttura per il
DNA; essi non eseguirono veri e propri esperimenti, ma intrapresero, piuttosto,
un esame razionale di tutti i dati allora noti sul DNA, cercando di
organizzarli in modo logico. Secondo il loro modello la molecola di DNA è
un'elica a filamento doppio, dalla forma di una scala a spirale. I due
«montanti» della scala sono costituiti da subunità ripetute di un gruppo
fosfato e dello zucchero deossiribosio a 5 atomi di carbonio. I «pioli» sono
costituiti da basi azotate appaiate (una purina si appaia con una pirimidina);
A può appaiarsi solo con T e G solo con C, e si chiamano complementari.
Le quattro basi sono le quattro «lettere» usate per scandire il messaggio
genetico. Le basi appaiate sono unite da legami a idrogeno.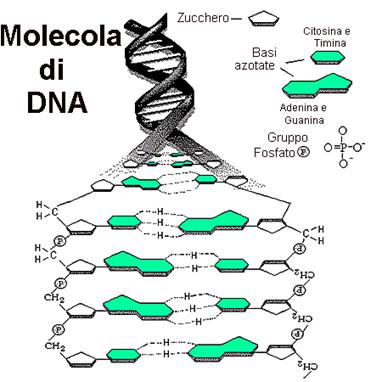
Duplicazione
Quando la molecola di DNA si duplica, i due filamenti si separano in seguito
alla rottura dei legami a idrogeno. Ogni filamento si comporta come uno stampo per
la formazione di un nuovo filamento complementare, utilizzando i nucleotidi
disponibili nella cellula. L'aggiunta di nucleotidi al nuovo filamento è
catalizzata dagli enzimi DNA-polimerasi. Nel processo di
duplicazione molti altri enzimi giocano un ruolo importante.
La duplicazione del DNA inizia a livello di una particolare sequenza di
nucleotidi sul cromosoma, che è il punto d'origine della duplicazione.
Questa procede in entrambe le direzioni, per mezzo delle due forcelle di
duplicazione che si spostano nelle due direzioni opposte. Durante la
duplicazione del DNA avviene l'azione proofreading delle DNA-polimerasi,
che fanno invertire la direzione di marcia quando si rende necessario rimuovere
quei nucleotidi che non si sono appaiati in modo corretto a quelli del
filamento stampo.
Grazie alle deduzioni sulla struttura a doppia elica del DNA elaborate da
Watson e Crick, venne universalmente accettato il ruolo del DNA come la
molecola che porta a trasmettere le informazioni genetiche. Con la
scoperta del complesso ed estremamente preciso meccanismo mediante il quale le
cellule duplicano il loro DNA era finalmente risolto il problema di come
l'informazione ereditaria venga fedelmente trasmessa da una cellula madre alla
cellula figlia, generazione dopo generazione.
LA TRADUZIONE DEL CODICE GENETICO
Un gene-un enzima
Ritornando un po' indietro nella storia notiamo come già negli anni '40 il
biologo Beadle notò l'importanza del legame che corre tra un gene ed il suo
corrispondente enzima. Beadle, insieme al chimico Tatum, nel 1941 incominciò ad
analizzare i mutanti di Neurospora (muffa rossa del pane) per
dimostrare, attraverso studi di mappatura genica, che a una certa mutazione
corrisponde la perdita di funzionalità di un certo enzima. Ma, visto che gli
enzimi sono composti da proteine e le proteine da catene polipeptidiche, allora
poterono dire che a un gene corrisponde una proteina o, più precisamente, una
catena polipeptidica (riassumendo con la formula un gene-una catena
polipeptidica).
DAL DNA ALLA PROTEINA: RUOLO
DELL'RNA
Il problema successivo divenne quello della traduzione: come faceva la
successione delle basi azotate a determinare la sequenza degli amminoacidi di
una proteina? La ricerca di una risposta a questo quesito condusse alla scoperta
dell'acido ribonucleico (RNA), una sostanza chimicamente simile
al DNA.
Come poi risultò, non uno ma tre tipi di RNA agiscono come intermediari nei
processi che, partendo dal DNA, portano alle proteine. Questi tre tipi si
distinguono in forma e funzionalità, ed agiscono l'uno dopo l'altro nella
traduzione. Si chiamano RNA
messaggero (mRNA), RNA
ribosomiale (rRNA) e RNA di trasporto (tRNA).
RNA messaggero (mRNA)
A questo punto della nostra spiegazione incominciamo a descrivere il primo: l'RNA
messaggero (mRNA). Le molecole di RNA messaggero sono copie
(trascrizioni) di sequenze nucleotidiche codificate del DNA. A differenza del
DNA, tuttavia, le molecole di RNA hanno in genere un solo filamento. Ogni nuova
molecola di mRNA viene copiata da uno dei due filamenti del DNA con lo stesso
principio che regole la duplicazione del DNA. L'aver identificato l'mRNA come
la copia di lavoro delle istruzioni genetiche non aveva ancora risolto il
problema di fondo. Le proteine contengono 20 amminoacidi differenti, ma il DNA
e l'RNA contengono ciascuno solo quattro diversi nucleotidi; in qualche modo
questi nucleotidi costituivano un codice genetico per gli amminoacidi.
Facendo un po' di calcoli su questi numeri si notò come l'unico modo in cui il
DNA avrebbe potuto codificare per gli amminoacidi era che un singolo
amminoacido dovesse essere determinato da tre nucleotidi in sequenza. Le 64
possibilità formate dalle composizioni dei nucleotidi vennero chiamate codoni.
Nirenberg e Matthaei fruttarono giusto l'RNA messaggero per
decifrare il codice genetico. Provarono ad inserire parti di una cellula che
permettessero di produrre proteine in una provetta dove inserirono degli RNA
artificiali che ripetevano uno stesso codone più volte. Un certo tipo di
codone, formato da tre nucleotidi, traduce per un certo tipo di amminoacidi che
legati insieme formano una proteina.
Ribosomi
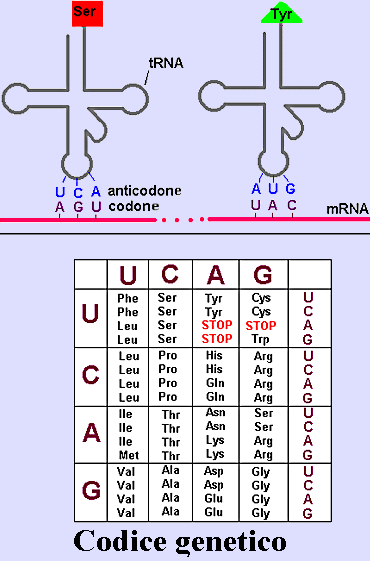 La sintesi proteica, o traduzione, avviene sui ribosomi.
Un ribosoma è formato da due subunità, una grossa e una piccola, ognuna
costituita da specifici RNA ribosomiali (rRNA) e da molecole
proteiche specifiche. Nella sintesi proteica è necessario anche un altro tipo
di molecole di RNA, cioè l'RNA di trasporto (tRNA) che ha forma
di trifoglio. Queste piccole molecole possono portare su un'estremità un
amminoacido e hanno una tripletta di basi, l'anticodone, sul «braccio» centrale
all'estremità opposta della molecola. Durante la sintesi, il tRNA funziona da
adattatore (quasi come un dizionario bilingue) mettendo in corrispondenza
ciascun codone dell'mRNA col relativo amminoacido; esiste almeno un tipo di
tRNA per ciascun tipo di amminoacido presente nella cellula.
La sintesi proteica, o traduzione, avviene sui ribosomi.
Un ribosoma è formato da due subunità, una grossa e una piccola, ognuna
costituita da specifici RNA ribosomiali (rRNA) e da molecole
proteiche specifiche. Nella sintesi proteica è necessario anche un altro tipo
di molecole di RNA, cioè l'RNA di trasporto (tRNA) che ha forma
di trifoglio. Queste piccole molecole possono portare su un'estremità un
amminoacido e hanno una tripletta di basi, l'anticodone, sul «braccio» centrale
all'estremità opposta della molecola. Durante la sintesi, il tRNA funziona da
adattatore (quasi come un dizionario bilingue) mettendo in corrispondenza
ciascun codone dell'mRNA col relativo amminoacido; esiste almeno un tipo di
tRNA per ciascun tipo di amminoacido presente nella cellula.
|
Privacy |
Articolo informazione
Commentare questo articolo:Non sei registratoDevi essere registrato per commentare ISCRIVITI |
Copiare il codice nella pagina web del tuo sito. |
Copyright InfTub.com 2025