|
|
| |
Compartimenti intracellulari e smistamento delle proteine
Cap
A differenza dei batteri che consistono generalmente di un unico compartimento intracellulare circondato da una membrana plasmatica, una cellula eucariota è suddivisa in compartimenti funzionalmente distinti, racchiusi da membrane. Ciascun compartimento, o organello, contiene la propria serie caratteristica di enzimi e di altre molecole. Le proteine conferiscono a ciascun compartimento le sue proprietà strutturali e funzionali, catalizzano le reazioni che hanno luogo in ciascun organello e trasportano selettivamente piccole molecole dentro e fuori dal suo interno, o lume.
Tutte le cellule eucariote hanno la stessa serie di base di organelli racchiusi da membrane. Quest'ultime hanno compartimenti chiusi separati da citosol.
Tutte le proteine presenti nella cel 616f53g lula iniziano con l'essere sintetizzate sui ribosomi nel citosol e il loro destino successivo dipende dalla loro sequenza amminica. Essa, infatti, può contenere segnali di smistamento che dirigono la loro consegna verso posizioni fuori dal citosol. La maggior parte delle proteine, però, non contiene nessun segnale di smistamento e resta, quindi, all'interno del citosol.
Esistono tre meccanismi principali tramite cui le proteine si muovono da un compartimento alla'ltro all'interno di una cellula:
Trasporto attraverso i pori: il traffico delle proteine fra il citosol e il nucleo avviene fra spazi equivalenti. I complessi dei pori nucleari servono da cancelli selettivi che trasportano attivamente macromolecole e complessi specifici.
Trasporto transmembrana: proteine traslocatrici, legate alla membrana trasportano direttamente proteine specifiche attraverso una membrana dal citosol in uno spazio distinto. La proteina trasportata, in genere, deve perdere i suoi ripiegamenti per passare come un serpente attraverso il trasfocatore.
Trasporto vescicolare: intermedi di trasporto racchiusi da membrana traghettano le proteine da un compartimento all'altro. Le vescicole e i frammenti di trasporto si caricano di molecole derivate dal lume di un compartimento quando si distaccano dalla sua membrana e scaricano il loro contenuto in un secondo compartimento fondendosi con quello.
Ciascuno dei tre meccanismi è guidato da segnali di smistamento nelle proteine trasportate che sono riconosciute da recettori proteici complementari. Esistono due tipi di segnali di smistamento nelle proteine. Un tipo si trova in un tratto continuo della sequenza di amminoacidi, lunga dai 15 ai 60 residui. Alcune di queste sequenze segnale sono rimosse dalla proteina una volta che lo smistamento è stato completato da una peptidasi di segnale. L'altro tipo consiste in una sequenza tridimensionale specifica di atomi sulla superficie della proteina che si forma sulla proteina stessa quando essa si ripiega.
Ciascuna sequenza segnale specifica una particolare destinazione nella cellula. Proteine destinate inizialmente nel reticolo endoplasmatico hanno una sequenza segnale al loro N-terminale. Molte di queste proteine passeranno poi all'apparato del Golgi, mentre quelle che restano nel reticolo endoplasmatico hanno una sequenza specifica di 4 amminoacidi al loro C-terminale. Entrambi i tipi di segnali di smistamento sono riconosciuti da recettori di smistamento che guidano le proteine alla loro destinazione appropriata.
![]() Componenti
cellulari:
Componenti
cellulari:
Il nucleo
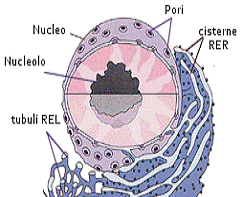 Il nucleo controlla la maggior parte delle attività della cellule e
gioco ruolo di primaria importanza della replicazione, nell'accrescimento e nel
differenziamento cellulare. Il nucleo contiene il Dna che risulta complessato
con proteine strutturali, gli istoni,
a costituire la cromatina.
Quando la cellula non è in divisione i filamenti di Dna che costituiscono i
diversi cromosomi
sono despiralizzati e formano un ammasso indistinto, mentre prima della
divisione cellulare la cromatina si addensa e i singoli cromosomi assumono
l'aspetto compatto con cui sono visibili al microscopio. Nel nucleo sono
inoltre contenuti uno o più nucleoli, particolari strutture in cui vengono
sintetizzati gli RNA ribosomiali (rRNA) e vengono assemblati i ribosomi.
Il nucleo controlla la maggior parte delle attività della cellule e
gioco ruolo di primaria importanza della replicazione, nell'accrescimento e nel
differenziamento cellulare. Il nucleo contiene il Dna che risulta complessato
con proteine strutturali, gli istoni,
a costituire la cromatina.
Quando la cellula non è in divisione i filamenti di Dna che costituiscono i
diversi cromosomi
sono despiralizzati e formano un ammasso indistinto, mentre prima della
divisione cellulare la cromatina si addensa e i singoli cromosomi assumono
l'aspetto compatto con cui sono visibili al microscopio. Nel nucleo sono
inoltre contenuti uno o più nucleoli, particolari strutture in cui vengono
sintetizzati gli RNA ribosomiali (rRNA) e vengono assemblati i ribosomi.
Il nucleo è circondato da una doppia membrana che lo separa dal citoplasma, la membrana nucleare, costellata di pori (i pori nucleari) che permettono scambi selettivi con il citoplasma.
La membrana nucleare interna contiene proteine specifiche che agiscono da siti di attacco per la cromatina, mentre la membrana nucleare esterna è in continuità con la membrana del reticolo endoplasmatico. Come quest'ultimo la membrana nucleare esterna è costellata da ribosomi impegnati nella sintesi proteica.
L'involucro nucleare di tutti gli eucarioti è, quindi, perforato da pori nucleari circondati da una struttura ad anello, detta complessi dei pori nucleari, formata da otto granuli di origine proteica. Nelle cellule animali si pensa che tali strutture siano costituite da più di 50 proteine diverse, chiamate nucleoporine.
Ciascun complesso contiene uno o più canali acquosi aperti attraverso i quali piccole molecole solubili in acqua possono diffondere passivamente. Ma poiché molte proteine sono molto grandi per poter passare attraverso questi pori per diffusione devono assemblarsi a recettori che le traghettano attivamente attraverso i pori nucleari. Per iniziare l'importazione nel nucleo i segnali di localizzazione (NLS) devono essere riconosciuti da recettori di importazione nucleare. Tali recettori sono proteine citosoliche solubili che si legano sia al segnale di localizzazione nucleare posto sulla proteina da trasportare che alle nucleoporine, alcune delle quali formano fibrille. Quest'ultime contengono numerosi ripetizioni di fenilanina e glicina chiamate per tale motivo ripetizioni FG. Tali ripetizioni servono da siti d'attacco per i recettori di importazione. Una volta nel nucleo tali recettori si dissociano e sono reimmersi nel citosol.
L'esportazione dal nucleo di grosse molecole avviene anch'essa attraverso i pori nucleari e dipende da un sistema di trasporto selettivo. Questo tipo di trasporto si basa su segnali di esportazione nucleare presenti sulle macromolecole e riconosciute da recettori di esportazione nucleare, i quali si legano sia alle nucleoporine che al segnale per guidare il carico attraverso il poro nucleare in modo tale che arrivi nel citosol.
Alcune proteine, però, contengono sia segnali di importazione che di esportazione, tali proteine vanno continuamente avanti e dietro fra il nucleo e il citosol. La localizzazione di queste proteine navetta è determinata dalle velocità relative di importazione e di esportazione. Infatti se la velocità di importazione supera quella di esportazione, la proteina si troverà soprattutto nel nucleo, viceversa nel citosol.
I mitocondri
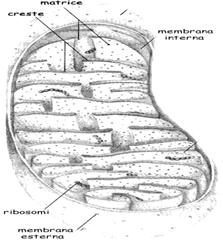 I mitocondri sono organelli delimitati da una membrana doppia che
delimitano due compartimenti: lo spazio della
matrice interno e lo spazio
intermembrana. Tali compartimenti sono formati da due
membrane concentriche: una interna che presenta numerose pieghe, dette creste e
racchiude lo spazio della matrice e una esterna liscia che è in continuità con
il citosol.
I mitocondri sono organelli delimitati da una membrana doppia che
delimitano due compartimenti: lo spazio della
matrice interno e lo spazio
intermembrana. Tali compartimenti sono formati da due
membrane concentriche: una interna che presenta numerose pieghe, dette creste e
racchiude lo spazio della matrice e una esterna liscia che è in continuità con
il citosol.
I mitocondri sono organelli di semiautonomi: possiedono un proprio DNA (circolare), ribosomi simili a quelli batterici (con i quali producono alcune proteine) e si dividono per scissione binaria.
Secondo la teoria dell'endosimbiosi sarebbero i discendenti di primitive cellule procarioti è che avrebbero instaurato una relazione simbiotica con l'antenato della cellule eucariote. Possono essere considerati le centrali energetiche delle cellule: sono infatti la sede della respirazione cellulare, processi in cui le sostanze organiche, in presenza di ossigeno, vengono demolite a CO2 e H2O, liberando energia che viene esportata per sintetizzare ATP, necessario per le attività cellulari.
La traslocazione delle proteine attraverso la membrana dei mitocondri è mediata da complessi proteici come il complesso tom, il quale svolge la sua funzione attraverso la membrana esterna ed è necessario per l'importazione di tutte le proteine mitocondriali e due complessi tim, i quali svolgono la loro funzione attraverso la membrana interna. L'energia richiesta è fornita dall'idrolisi di ATP.
I perossisomi
I perossisomi differiscono dai mitocondri e ai cloroplasti. La differenza più evidente è che i perossisomi sono circondati da un'unica membrana e non contengono né DNA né ribosomi. Come i mitocondri e i cloroplasti acquisiscono le loro proteine dal citosol, ma poiché sono privi di genoma devono importare tutte le proteine. In questo aspetto somigliano al reticolo endoplasmatico che pur non avendo un genoma proprio sono capaci di sopravvivere e di replicarsi. I perossisomi si trovano in tutte le cellule eucariote e contengono enzimi ossidativi, il loro nome deriva proprio dal fatto che contengono uno o più enzimi che usano ossigeno molecolare per rimuovere atomi di idrogeno in una reazione ossidativa che produce acqua ossigenata (H2O2). Una funzione importante delle reazioni ossidative è quella di eliminare acidi grassi mediante un processo noto come β eliminazione.
Reticolo endoplasmatico e apparato di Golgi
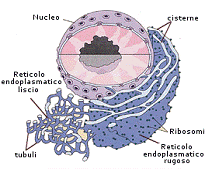 Il reticolo
endoplasmatico (RE) è il sistema
di membrane più esteso della cellula ed è costituito da una fitta rete di cisterne
e tubuli tra loro interconnessi che parte ed è collegato alla
membrana nucleare. La membrana dell'RE forma un foglio continuo che racchiude
uno spazio interno chiamato lume dell'RE. La membrana dell'RE separa il
lume dell'RE dal citosol e media il trasferimento selettivo di molecole fra
questi due compartimenti.
Il reticolo
endoplasmatico (RE) è il sistema
di membrane più esteso della cellula ed è costituito da una fitta rete di cisterne
e tubuli tra loro interconnessi che parte ed è collegato alla
membrana nucleare. La membrana dell'RE forma un foglio continuo che racchiude
uno spazio interno chiamato lume dell'RE. La membrana dell'RE separa il
lume dell'RE dal citosol e media il trasferimento selettivo di molecole fra
questi due compartimenti.
La membrana dell'RE è il sito di produzione di tutte le proteine transmembrana e di tutti i lipidi. L'ER cattura le proteine dal citosol mentre queste vengono sintetizzate. Queste proteine possono essere di due tipi:
Proteine transmembrana: traslocate attraverso la membrana dell'RE e vi restano immerse in essa.
Proteine solubili in soluzioni acquose: sono completamente traslocate attraverso la membrana dell'RE e sono poi rilasciate nel lume dell'RE.
Nelle cellule di mammifero, l'importazione di proteine nell'RE comincia prima che la catena polipeptidica sia completamente sintetizzata. Questo tipo di importazione è detta contraduzionale. Essa si distingue completamente dall'importazione di proteine che avviene nei cloroplasti e nei mitocondri, in quanto tale importazione è post-traduzionale.
I ribosomi che rivestono la superficie dell'RE creano regioni chiamate reticolo endoplasmatico ruvido o RE ruvido. Sono quindi presenti due tipi di ribosomi nel citosol, quelli legati alla membrana, impegnati nella sintesi delle proteine che stanno traslocando nell'RE, e i ribosomi liberi, non attaccati a nessuna membrana, i quali sintetizzano tutte le altre proteine codificate dal genoma.
Le regioni dell'RE prive di ribosomi costituiscono il reticolo endoplasmatico liscio, o RE liscio. Tale reticolo ha funzioni importanti come ad esempio partecipa alla catalisi per la detossificazione da farmaci o veleni o di componenti dannosi prodotti dal metabolismo.
|
Privacy |
Articolo informazione
Commentare questo articolo:Non sei registratoDevi essere registrato per commentare ISCRIVITI |
Copiare il codice nella pagina web del tuo sito. |
Copyright InfTub.com 2026