|
|
| |
Neurofisiologia generale
Ogni cellula è costituita da membrana plasmatici, e quest'ultima è costituita da lipidi che contengono: colina, colesterolo e glicolipidi; inoltre è costituita da fosfolipidi composti da acidi grassi saturi e uno insaturo.
La fluidità e la rigidità della membrana sono regolate dalla presenza di catene insature e dal colesterolo.
Le proteine sono rappresentate da varie molecole con funzioni diverse, enzimi, recettori, antigeni, interagiscono con altre cellule presenti nel liquido extracellulare. Sono:
-proteine intrinseche o integrali, nel doppio strato lipidico
-proteine estrinseche o periferiche, nella superficie della membrana
La disposizione delle proteine nel doppio strato lipidico rende la membrana cellulare paragonabile ad un mosaico fluido. Alcune di queste proteine sono glicoproteine costituite da catene laterali glucidiche e sono situate sul lato extracellulare. In questa zona si trovano le molecole-recettore, sorta di antenne che sentono un messaggio chimico che arriva dalla cellula. Dopo che il messaggio viene trasferito sul lato interno della membrana si attiva un'altra serie di proteine, proteine-canale, e la risposta al messaggio proviene dal nucleo della cellula. Caratteristiche comuni a recettori ed enzimi sono la sensibilità e la specificità per il ligando.
Le membrane biologiche sono barriere di permeabilità che impediscono a gran parte delle molecole presenti nei sistemi viventi solubili in acqua di diffondere. Le membrane separano i vari componenti e scelgono le modalità e le tempistiche del passaggio di molecole e ioni, con meccanismi diversi di trasporto:
trasporto senza attraversamento della membrana passaggio di sostanze da un lato all'altro della membrana per mezzo di vescicole
-endocitosi, meccanismo attraverso il quale la cellula è in grado di inglobare materiale presente all'esterno per portarlo al suo interno. Avviene nella superficie esterna dove si trovano le molecole ancorate che raccolgono il materiale da portare all'interno mentre nella superficie interna vi sono strutture filamentose proteiche
-esocitosi, meccanismo che permette la fuoriuscita di sostanze dalla cellula
trasporto con attraversamento della membrana è formato da:
-trasporto in forma libera in cui la sostanza passa attraverso la membrana senza l'intervento di molecole trasportatrici
-trasporti mediati il cui passaggio avviene per mezzo di molecole trasportatrici, le caratteristiche sono:
velocità, sostanze trasportate presentano una velocità di trasferimento maggiore rispetto a quella di semplice diffusione
cinetica di saturazione, la velocità di trasporto di una sostanza aumenta fino a che l'incremento diventa nullo e la velocità invariata
inibizione da molecole non simili, un composto dissimile si può legare al trasportatore riducendone l'affinità per il normale substrato. Il trasporto facilitato non dipende dall'energia metabolica
-trasporti passivi senza consumo di energia e senza l'intervento di molecole trasportatrici (diff libera) o con il loro intervento (diff facilitata)
-trasporti attivi con consumo di energia derivata dall'idrolisi di legami chimici. Vengono chiamati primari quando il consumo di energia è direttamente coinvolto nel trasporto della sostanza. Agisce contro gradiente di concentrazione e/o elettrochimico, per la cui attivazione è necessario un diretto consumo di ATP, e legato al metabolismo energetico cellulare. La sua esistenza dipende dalla presenza di:
substrati proteici di trasporto
cambio conformazionale delle proteine trasportatrici
La possibilità che si verifiche un trasporto attivo è determinata dalla presenza di proteine di trasporto specifiche che si alternano in due conformazioni:-i siti di legame cono rivolti al liquido extracellulare
-i siti di legame sono rivolti al citosol
Le correnti ioniche attraversano la membrana plasmatici per mezzo di proteine intrinseche di membrana che si dispongono a formare dei pori chiamati "CANALI". I canali, ad eccezione di quelli che determinano il potenziale di riposo, possono fluttuare tra uno stato di apertura e uno di chiusura. Quando il canale è attivo, la frequenza dello stato di apertura predomina sullo stato di chiusura, aumentando il flusso di correnti ioniche tra interno ed esterno della cellula.
Classificazione e struttura molecolare in funzione delle modalità operative possono essere distinti in:
-canali ionici passivi e aperti da voltaggio (voltaggio - dipendenti), al quale appartengono i canali legati al mantenimento del potenziale di riposo. I canali del sodio voltaggio - dipendenti, ma anche quelli del potassio e del calcio, si aprono in risposta alla depolarizzazione di membrana, lasciando passare gli ioni durante la fase di salita del potenziale d'azione. Spontaneamente però si inattivano.
-canali ionici aperti da ligandi (ligando - operante), sono canali il cui stato di apertura è regolato da un segnale chimico. A questo gruppo appartengono sia canali attivi da proteine traduzione, sia canali recettori che riconoscono il ligando specifico e aprono un varco attraverso cui passano gli ioni
-canali ionici delle gap junction, sono canali presenti a livello di tessuti in cui la comunicazione cellula - cellula deve essere sincrona e particolarmente veloce. Questo tipo di canale nel sistema nervoso è diffuso nelle sinapsi elettriche. Le giunzioni comunicanti sono costituite da due strutture, una che appartiene alla cellula postsinaptica e una a quella presinaptica. Interagiscono tra loro formando un canale che pone in comunicazione il citoplasma delle due cellule contigue.
L'accesso o la chiusura di un canale può essere controllata da stimoli diversi. Ci sono canali la cui apertura è dominata da processi di fosforilazione e defosforilazione che avvengono grazie al trasferimento di un fosfato ad alta energia su residui amminoacidi. Altri canali, invece, possono essere attivati da pressioni meccaniche e dallo stiramento di membrana. Anche i processi di inattivazione sono molteplici. Per esempio in alcuni canali del calcio voltaggio - dipendenti l'aumento della concentrazione citoplasmatica dello ione, dovuta alla apertura del canale in risposta alla depolarizzazione, funge da elemento inattivante per lo stesso (omologa) o per altri tipi di canali (eterologa).
La funzionalità dei sistemi cellulari, come la capacità di un neurone di trasportare informazioni proprie è determinato dall'esistenza di una differenza di potenziale tra i due versanti della membrana citoplasmatica.
La presenza di un potenziale di membrana (potenziale a riposo) e la possibilità di modificarlo in potenziale d'azione in risposta di precisi eventi, costituiscono per la cellula uno dei più formidabili ed evoluti strumenti di trasmissione dei segnali intercellulari.
Le variazioni transitorie del potenziale di membrana rispetto al suo normale valore sono la conseguenza di modificazioni del flusso di correnti totali che entrano ed escono dalla cellula. Il flusso è regolato dai canali ionici:
Canale ionico passivo (sempre aperto) determinazione del potenziale di riposo;
Canale attivo determinate per l'insorgenza del potenziale d'azione.
Analizzando la composizione chimica dentro la cellula si ha molta concentrazione di K mentre all'esterno della cellula si ha maggiore concentrazione di Na. Nella maggior parte delle cellule si osserva un'eccedenza di cariche negative sulla faccia interna della membrana mentre dalla parte esterna vi è un eccesso di cariche positive. Nei neuroni il potenziale di membrana a riposo varia tra -60 e -70 mV, per convenienza si assume pari a zero il valore del potenziale all'esterno, per cui il potenziale di membrana (Vm) sarà:
Vm = Vi - Ve Vi = potenziale esistente all'interno della cellula
Ve = potenziale all'esterno.
Ogni situazione che porta alla diminuzione di Vm, cioè a una diminuzione della differenza di potenziale ai lati della membrana, prende il nome di depolarizzazione; si chiamerà iperpolarizzazione la situazione inversa.
Equazione di Nernst il potenziale di equilibrio dipende dalla quantità di ioni da una parte e dall'altra.
E' dovuto dalla presenza nella membrana plasmatica di canali ionici passivi per gli ioni K . Il k è concentato nel citoplasma e tende a migrare verso l'ambiente extra essendo la membrana scarsamente permeabile agli anioni per la scarsità di canali, il flusso continuo di ioni K determinerà un eccesso di cariche positive (cationi) all'esterno della cellula e, un eccesso di anioni e un eccesso di cariche negative all'interno. Questa diseguale distribuzione di ioni è responsabile della comparsa di cariche positive a ridosso del versante esterno della membrana e di cariche negative all'interno. Il movimento di K è dato dalla sottoposizione a due forze, una di repulsione e l'altra di attrazione elettrostatica dovuta al suo movimento. Quindi su ogni ione K vanno ad agire due forze una di natura chimica e uno di natura elettrica. Il gradiente di concentrazione tende a spingere K verso l'esterno della cellula, mentre quello elettrico darà origine ad una differenza di potenziale elettrico che tende a spingere il catione all'interno.
E' costituito da una variazione rapida del potenziale di membrana, con ritorno veloce al valore precedente. La sua forma e durata dipendono dal tipo di tessuto eccitabile. Normalmente i potenziali d'azione sono molto veloci nei neuroni e nel muscolo striato scheletrico. Attraverso i potenziali d'azione le cellule nervose inviano segnali ad atre cellule nervose o muscolari in cui innescano la contrazione. Nella maggior parte dei neuroni, portando il potenziale a un valore soglia pari a -55 mV (potenziale di soglia), si instaura un fenomeno elettrico chiamato potenziale d'azione, risposta così ampia che porta ad un'inversione di polarità che si propaga lungo la fibra. L'ampiezza e la forma del potenziale d'azione rimangono costanti anche se si continua a stimolare a maggior intensità e con un tempo più lungo di applicazione. In parole più semplici, non si ha gradualità nella risposta: uno stimolo o è capace di provocare l'intera manifestazione (soprasoglia) oppure non lo è (sottosoglia). Per queste ragioni il potenziale d'azione si definisce "o tutto o nulla" . Una volta che la membrana ha raggiunto il valore soglia, inizia una fase di rapida depolarizzazione che supera il limite dello zero (over - shoot), consentendo all'apice del potenziale d'azione di raggiungere un valore positivo intorno a 35 mV. Abbastanza rapidamente il potenziale di membrana ritorna verso i valori di riposo. Dopo la fase di depolarizzazione si osserva una fase di iperpolarizzazione che dura alcuni secondi.
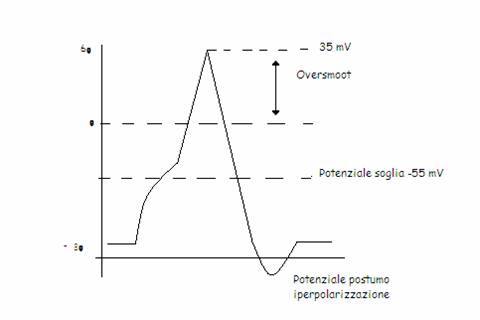
Basi ioniche
Durante il potenziale d'azione si hanno variazioni precise della conduttanze (g) di alcune specie ioniche. In particolare, si ha un aumento sequenziale della conduttanza del sodio (gNa) e del potassio (gK), conseguente alla variazione dello stato funzionale (apertura) di canali voltaggio - dipendenti selettivi per queste due specie ioniche.
Durante la depolarizzazione, l'apertura massima dei canali sodio voltaggio - dipendenti, permette ai due gradienti passivi di determinare il rapido ingresso di ioni Na all'interno della cellula, causando uno spostamento del potenziale di membrana da valori vicini al potenziale di equilibrio del K (-70 mV) verso valori più prossimi al potenziale di equilibrio del Na (55 mV). In realtà non si raggiunge questo valore ma il picco del potenziale d'azione si ferma a 35 mV. A questo punto i canali del sodio si disattivano, mentre i canali del K raggiungono e mantengono il massimo stadio di apertura. Questo stato di inversione del potenziale e il mantenimento dello stato aperto dei canali di rettifica del K permettono ad ambedue i gradienti di cooperare alla fuoriuscita del K e quindi a un ritorno del potenziale su valori prossimi del potenziale di riposo.
Propagazione del potenziale elettrico "tutto o nulla"
La velocità di conduzione dipende sia dalla proprietà fisica del segnale elettrico sia a quelle legate alle caratteristiche fisica della fibra nervosa. La propagazione degli eventi graduati è influenzata dalla resistenza passiva di membrana e quindi dalla sua composizione (densità e distribuzione di canali come vie a bassa resistenza ai flussi di corrente). Queste proprietà tipiche di ogni membrana sono rappresentate dalla costante di spazio. Il segnale si propaga longitudinalmente nel citoplasma, incontrando resistenze e conduttanze diverse rispetto a quella della membrana. La combinazione di questi fattori determina la velocità di propagazione e l'entità del decremento del potenziale elettrico, caratteristiche definite dalla costante tempo. La corrente entra nella membrana plasmatica e si propaga nel citoplasma, fino a incontrare canali di membrana a bassa resistenza, per poi uscire dalla cellula e chiudere il circuito locale, una corrente in entrata o in uscita determinerà un altro circuito locale e così via, con andamento ad anelli sequenziali. La velocità di tale conduzione dipende dalle proprietà elettriche del citoplasma e della membrana: più è grande il diametro della fibra, maggiore è la velocità.
Propagazione del potenziale d'azione
Se il circuito locale ha un'intensità sufficiente e incontra zone particolari della membrana, può causare l'insorgenza del potenziale d'azione. I potenziali d'azione sono condotti lungo la fibra nervosa per trasferire informazioni ad altri neuroni, a cellule somatiche e a fibre muscolari, che a loro volta sono capaci di condurre il segnale all'interno della fibra stessa. Il potenziale d'azione si genera in seguito alla propagazione elettrotonica di circuiti depolarizzanti locali. Questa differenza di potenziale genera flussi locali di correnti ioniche che portano cariche positive all'interno della cellula e sottraggono cariche positive all'esterno. Quando la depolarizzazione raggiunge il valore soglia e viene attivato un adeguato numero di canali del sodio voltaggio-dipendenti, allora si genera il potenziale d'azione. La propagazione del potenziale d'azione lungo tutta la fibra è possibile solo grazie alla generazione di nuovi potenziali d'azione che si innescano autonomamente nelle zone contigue a quelle in cui è stato raggiunto il potenziale di soglia. La direzione è sempre una: quella che tende ad allontanarsi dal punto di insorgenza del potenziale d'azione e sarà unidirezionale. Negli assoni più lunghi la guaina mielinica presenta interruzioni ogni 1-2 mm, nodi di Ranvier. La guaina determina un aumento della velocità di conduzione del segnale. Attraverso la guaina si crea un aumento della resistenza della membrana. L'aumento della velocità di conduzione che si ha in presenza della guaina è dovuto ad un aumento della costante di spazio e ad una riduzione della capacità di membrana. Grazie all'isolamento mielinico viene perso meno segnale e il risultato è un aumento della velocità di conduzione. I potenziali d'azione si propagano saltando da un nodo all'altro procedendo più velocemente nelle fibre mieliniche: conduzione saltatoria.
Classificazione delle fibre nervose a trasmissione sinaptica
-pirenoforo: corpo cellulare che contiene il nucleo e i vari organuli cellulari
-assone: lunga struttura tubulare che, partendo dal soma con il cono di emergenza si prolunga fino alla cellula bersaglio, il loro compito è quello di condurre gli impulsi lontano dal corpo cellulare. Possono essere fibre mieliniche o fibre amieliniche. Nel SNP la guaina mielinica è costituita dalle cellule di origine gliale, dette cellule di Schwann, quelt'ultime si avvolgono agli assoni in modo da creare una struttura isolante e inoltre queste guaine mieliniche separano le cellule di Schwann creando i nodi di Ranvier. L'assone termina con la membrana giunzionale o bottone terminale
-dendriti: costituiscono la chioma del corpo cellulare e terminano con assoni provenienti da altri neuroni, sono deputati a ricevere i segnali e, solo nella sinapsi dendrido-dendritica trasmette le informazioni mediante un proprio bottone sinaptico.
In base alla funzione possono essere neuroni afferenti, che portano informazioni da tutto il corpo, oppure neuroni efferenti che portano l'info dall'encefalo al midollo spinale. Il trasporto assonale si avvale di strutture citoscheletriche deputate a trasportare proteine e vescicole dal soma verso il bottone sinaptico. Le principali proteine trasportate sono la chinesina e la dineina. Questi trasporti monodirezionali possono svolgersi:
-dal soma verso il bottone terminale (trasporto anterogrado);
-dal bottone terminale verso il pirenoforo (trasporto retrogrado)
Grazie alla chinesina e alla dineina, proteine "carrello" che sono distribuite e ancorate lungo i binari microtubulari.
Classificazione delle fibre nervose
La capacità di conduzione del segnale elettrico è dato dalle fibre mieliniche, più veloci, e amieliniche, più lente.
Una più accurata classificazione, basata sulla velocità di conduzione del segnale elettrico, prevede una suddivisione delle fibre in:
-fibre di gruppo A, mieliniche dei nervi periferici, a loro volta si suddividono in:
a con velocità di conduzione di 80-120 m/s
b con velocità di conduzione di 35-80 m/s
d con velocità di conduzione di 5-30 m/s
Inoltre le fibre che provengono dai muscoli sono state classificate in:
-fibre di gruppo I conduzione delle fibre Aa, e alloro volta suddivise in fibre Ia (fibre afferenti) e fibre Ib (provengono da recettori tendinei sensoriali
-fibre di gruppo II con velocità pari a quelle delle Ab
-fibre di gruppo III riferite alle fibre di tipo Ad
-fibre di gruppo B, appartengono alle fibre mieliniche del SNA
-fibre di gruppo C, fibre amieliniche a lenta conduzione.
Trasmissione sinaptica
La struttura cellulare del SN è costituita da neuroni, i quali comunicano tra loro e con i loro effettori attraverso il rilascio di sostanze chimiche chiamate neurotrasmettitori.
sinapsi elettrica: la natura fisica dell'informazione consiste nel trasferimento corretto della corrente elettrica per mezzo di ioni che attraversano la membrana di due cellule contigue uniti da canali proteici a bassissima resistenza, definiti giunzioni comunicanti o gap junction. Nella giunzione comunicante la corrente scorre da un neurone all'altro, come in continuità citoplasmatica, senza ritardo sinaptico e il segnale può decorrere sia in direzione unica (sinapsi elettrica rettificata) che bidirezionalmente. Questo tipo di trasmissione è detto elettrico e può avvenire in seguito alla depolarizzazione con aumento del calcio intracellulare.
sinapsi chimiche e loro classificazione: si caratterizzano per l'utilizzo di neurotrasmettitori, come vettori per il trasferimento delle informazioni da un neurone all'altro. La sinapsi è formata da tre parti principali:
-membrana presinaptica membrana terminale dell'assone
-spazio giunzionale lungo 16-30 mm, situato tra due neuroni, è detto anche fessura sinaptica
-membrana postsinaptica membrana cellulare di un altro neurone specializzata nella trasmissione dell'info
Le sinapsi possono essere differenziate anche in base alla componente postsinaptica che può essere rappresentata da un altro assone, da un dendrite o dal corpo di un altro neurone. Anche la forma delle vescicole contenenti il neurotrasmettitore può essere utilizzata come elemento distintivo, esistono due tipi di sinapsi:
-tipo I di Gray: formato da proteine disposte assimetricamente tra la membrane pre-postsinaptica che hanno trasmissione di tipo eccitatorio;
-tipo II di Gray: formato da vescicole ovoidali che contengono un neurotrasmettitore con azione inibitoria e si trovano tra la membrana pre e postsinaptica distribuite simmetricamente.
Il neurone che trasmette è detto membrana presinaptica, libera il neurotrasmettitore e il ricevente è la membrana postsinaptica. Tra le due membrane c'è uno spazio intersinaptico dove sono presenti proteine che regolano il mantenimento della giunzione e il metabolismo del neurotrasmettitore liberato. La terminazione sinaptica contiene diverse vescicole sinaptiche che possono contenere diversi neurotrasmettitori (acetilcoline, acido glutaminico, noradrenalina).
Meccanismo d'azione delle sinapsi
Il neurone presinaptico riceve un segnale depolarizzante che raggiunge la soglia a livello del cono d'emergenza, si forma così il potenziale d'azione che percorre l'assone arrivando fino al bottone sinaptico. La presenza di Ca permette al potenziale d'azione di aprire un conduzione dove vi è l'aumento della concentrazione intracellulare.
Schematizzazione:
-l'arrivo del potenziale d'azione al bottone sinaptico causa una depolarizzazione della terminazione presinaptica con conseguente apertura dei canali Ca e l'ingresso di Ca2 nella terminazione
-poi si ha la fusione della membrana delle vescicole con quella delle zone attive e la liberazione delle vescicole in tali zone
-il neurotrasmettitore diffonde fino ad incontrare i recettori posti sulla membrana postsinaptica
-il mediatore chimico legandosi hai recettori provoca un aumento della conduzione
Un ruolo importante nella trasmissione sinaptica, viene fatto dal Ca che ha un ruolo fondamentale nell'attuazione dell'intero sistema. In condizione di riposo (-70mV) i canali del Ca sono chiusi e alcune vescicole sono già localizzate nelle zone attive, ancorate alla membrana presinaptica tramite proteine di fusione. L'ancoraggio avviene grazie all'interazione omofilica tra la porzione membranaria e la porzione vescicolare delle proteine di fusione. La maggior parte delle altre vescicole si trova ancorata ai filamenti di actina del citoscheletro sopra le zone attive. Quando il potenziale d'azione arriva a -40mV si aprono i canali del Ca permettendo al catione di invadere le zone attive e di portare le vescicole verso le zone attive per favorire lo svuotamento. Avviene il rilascio del neurotrasmettitore in pochi millisecondi grazie al poro-canale, formato da proteine di fusione. Le molecole di neurortasmettitori si trovano disciolte nel liquido dello spazio sinaptico e le vescicole sono chiuse e trasportate lontano dalle zone attive per essere riempite.
I recettori
Qualunque variazione di energia proveniente dall'ambiente sia interno sia esterno all'organismo è captata da cellule specializzate, i recettori, e trasformata in una variazione di potenziale locale che, se le condizioni lo permettono, può generare una scarica di potenziale d'azione. Questi impulsi percorrono le fibre afferenti e arrivano al SNC dove vengono elaborati. I recettori si dividono in due grandi categorie:
-esterocettori: ricevono informazioni dall'ambiente esterno, comprendono il sottogruppo dei telecettori, cellule trasduttive che sono in grado di captare stimoli lontani
-intercettori: si dividono in propriocettori che registrano modificazioni energetiche che si verificano in altri distretti (es. stiramento muscolare)
classificazione in base alla forma di energia si distinguono in:
-chemiocettori: riconoscono con alta discriminazione specifiche molecole chimiche, sono in grado di dare informazioni specifiche sulle caratteristiche organolettiche dell'ambiente esterno o sullo stato chimico-fisico di quello interno
-meccanocettori: capaci di distinguere variazioni di energia meccanica come la pressione esercitata su di essi direttamente dall'esterno
-termocettori: captano le variazioni termiche dell'ambiente interno ed esterno
-recettori del dolore: possono essere considerati una sorta di chemiocetttori sensibili alle sostanze liberate dalla necrosi tissutale.
classificazione in base alle caratteristiche anatomo-funzionali si dividono in due tipologie:
-terminazione nervosa libera: porzione terminale non mielinizzata di un neurone sensitivo. Questo tipo di recettori è definito del I tipo. I recettori che contraggono sinapsi con una fibra afferente sensitiva si definiscono recettori del II tipo, infine, le cellule collegate alle fibre afferenti tramite un interneurone si definiscono recettori del III tipo
-cellule specializzate in contatto sinaptico con le fibre afferenti
i recettori in base al grado di adattamento si distinguono in:
-recettori tonici: danno una scarica di impulsi a frequenza costante per tutta la durata dello stimolo
-recettori fasici: a rapido adattamento, la cui scarica inizia in seguito alla stimolo e diminuisce subito dopo se lo stimolo è costante.
Riflessi spinali
Il riflesso è una risposta a stimolazioni esterne o interne diretta modificare lo stato di contrazione muscolare o di secrezione ghiandolare. Gli elementi costitutivi dei riflessi sono:
-recettori
-vie afferenti
-centro nervoso
-vie efferenti
-effettore
Quando uno stimolo colpisce una determinata area, vengono attivati i recettori e le vie afferenti. Gli impulsi raggiungono il midollo spinale ed eccitano direttamente o indirettamente tramite interneuroni, le fibre efferenti. Il tempo per scatenare la risposta dipende dalla velocità di conduzione delle fibre nervose e dal numero delle sinapsi centrali.
classificazione dei riflessi a livello spinale si riconoscono due grandi famiglie di riflessi:
-riflessi propriocettivi: originano dalla stimolazione di propriocettori muscolari e provocano la contrazione dei muscoli corrispondenti. L'intensità dello stimolo provoca l'aumento dell'intensità della contrazione muscolare. Il significato di questi riflessi è quello di mantenere la postura, controllare la lunghezza e la forza muscolare e facilitare la locomozione. Di questi riflessi fanno parte:
-riflesso miotatico fasico: forza provocata su un segmento del corpo e provoca l'allungamento brusco del muscolo. L'allungamento stimola i fusi neuromuscolari che sono all'interno del muscolo, dove si provoca una scarica di impulsi che raggiungono il midollo spinale e queste fibre si connettono con i motoneuroni e ne provocano un'eccitazione, ovvero si ha una contrazione muscolare.
-riflesso miotatico tonico: è attivato dalla forza di gravità che tende ad allungare i muscoli con stimolo e risposta continua.
-riflesso inverso da stiramento: frequenza di scarico proporzionale alla tensione del muscolo, le scariche raggiungono il midollo spinale attivando interneuroni che sono nel muscolo, e questi recettori aumentano la tensione e provocano un riflesso inverso da stiramento che riduce la forza muscolare.
-riflessi esterocettivi: originano dalla stimolazione della cute o delle mucose e danno luogo a risposte atte a proteggere la zona stimolata. Esempi sono i riflessi addominali, plantare, cutaneo.
|
Privacy |
Articolo informazione
Commentare questo articolo:Non sei registratoDevi essere registrato per commentare ISCRIVITI |
Copiare il codice nella pagina web del tuo sito. |
Copyright InfTub.com 2024